Eine ausführliche Buchbesprechung

Foto: Pexels.com / Alina Rossoshanska
In dem Buch „Schöpfung und Evolution? Drei Wissenschaftler. Drei Positionen. Eine Debatte“ diskutieren drei Autoren verschiedene Interpretationen von „Schöpfung“ und die Frage, ob Evolution und Schöpfung, Bibel und wissenschaftliche Erkenntnisse miteinander vereinbar sind. Lässt sich Schöpfung als Evolution denken oder ist die Bibel wörtlich zu verstehen? Lässt sich die kreationistische Idee einer wenige tausend Jahre jungen Welt auf empirische Befunde stützen? Vor allem: Wie gut belegt ist die Historizität des Evolutionsgeschehens und kann eine „Makroevolution“ natürlich ablaufen? Zum einen sagt die Diskussion manch Interessantes über die weltanschauliche Motivation der Autoren aus. Andererseits macht sich hinsichtlich der wissenschaftlichen Kompetenz von DROSSEL über SCHERER zu JUNKER ein deutliches Gefälle bemerkbar.
Die Autoren
Barbara DROSSEL ist Professorin für theoretische Physik an der TU Darmstadt. Sie forscht an der mathematischen Modellierung biologischer Systeme und an Grundfragen der Physik. Wenngleich sie einer Freikirche angehört (S. 347) und an „Schöpfung“ glaubt (S. 45), bricht sie eine Lanze für die akademische Evolutionstheorie.
DROSSEL distanziert sich klar von einem Junge-Erde-Kreationismus, weil dadurch der Glaube für naturwissenschaftlich gebildete Menschen intellektuell unannehmbar würde (S. 351). Auch den Kreationismus in seiner weiter gefassten Form des intelligenten Designs lehnt sie ab. Gleichwohl äußert sie den Wunsch, dass mehr junge Christen Physik oder Biologie studieren mögen, um mehr über Gottes großartige Schöpfung zu erfahren (S. 347). Je mehr Christen sich im akademischen Umfeld positionierten, desto wirkungsvoller, so DROSSEL, könnten sie atheistischen und naturalistischen Interpretationen der wissenschaftlichen Entdeckungen entgegentreten und das intellektuelle Klima beeinflussen. Dazu vertraut sie auf eine Stärkung durch Gott (S. 347)!
Reinhard JUNKER studierte Mathematik und Biologie für das Lehramt und bildete sich danach theologisch weiter. Später promovierte er zum Doktor der Theologie über eine kritische Beurteilung „theistischer Evolution“. Zwischen 1985 und 2021 arbeitete er bei der kreationistischen Studiengemeinschaft WORT UND WISSEN. Als ihr langjähriger Geschäftsführer sieht er sich einer unverfälschten Verkündigung der biblischen Botschaft verpflichtet, die nicht an „weltliche Philosophien angepasst“ sei (vgl. S. 288).
Als vermeintlich wissenschaftliche Alternative zur Evolutionstheorie vertritt JUNKER das kreationistische Grundtypenkonzept (S. 159) sowie den Design-Ansatz (vgl. dazu S. 125, 151, 306). Nach seiner Überzeugung sei dieser keine Theologie (S. 134). Zugleich glaubt er als Repräsentant eines Junge-Erde-Kreationismus (S. 288; S. 378 f.) nicht an die langen geologischen Zeiträume und lehnt „Makroevolution“ als natürlichen sowie auch als göttlich „gesteuerten“ Vorgang ab.
Um in der Tradition des Scientific Creationism eine junge Erde auch mit wissenschaftlichem Anspruch vertreten zu können, nennt er Einzelbefunde, die vermeintlich nicht zu den etablierten Datierungen passen. Losgelöst von der gesamtwissenschaftlichen Beleglage muss er dabei so viel astro- und kernphysikalisches, geologisches und kosmologisches Hintergrundwissen sowie die umfassende Evolutionshistorie ablehnen, dass vom etablierten Theoriennetzwerk nur noch ein Torso übrig bleibt. Er ignoriert somit das wichtige Wissenschaftskriterium der externen Konsistenz (VOLLMER 1995, S. 101).
Siegfried SCHERER nimmt im Meinungsspektrum des Autorenkollektivs eine Art Mittelposition ein. Er studierte Biologie, Chemie und Physik. Bis 2021 leitete er als Professor an der TU München einen mikrobiologischen Lehrstuhl. 1988 wurde er in den Leitungskreis von WORT UND WISSEN gewählt und war viele Jahre lang ihr ehrenamtlicher Vorsitzender. Lange galt er, gemeinsam mit JUNKER, als Wortführer des Junge-Erde-Kreationismus. Im Gegensatz zu JUNKER löste er sich jedoch vor etwa zwei Jahrzehnten davon (vgl. S. 180).
In den 1990er-Jahren lernte SCHERER die Intelligent-Design-Bewegung in den USA kennen und wurde Fellow des Discovery Institutes in Seattle. Dieses Institut ist ein Unternehmen des politischen Lobbyismus; Taede A. SMEDES stuft die Bewegung als politische Ideologie ein (JANOWSKI et al. 2010, S. 14). Später gab SCHERER seinen Fellow-Status aufgrund der zu politischen Orientierung wieder auf, weil sie einem wissenschaftlichen Diskurs nicht förderlich sei (S. 180 f.). Das heißt aber nicht, dass er sich vollends von Intelligent Design gelöst hätte, in mancherlei Hinsicht ist er nach wie vor dessen Anhänger (vgl. S. 253 f.). Seit 2019 unterstützt er das österreichische Zentrum für Biokomplexität und Naturteleologie (Leiter: Günter BECHLY) (S. 182), einen deutschsprachigen Ableger des Discovery Institutes, der sich Intelligent Design verpflichtet sieht.
Gliederung
Das Buch beginnt mit einer Darstellung der Positionen der drei Autoren Barbara DROSSEL, Reinhard JUNKER und Siegfried SCHERER. Es schließt sich eine konstruktiv-kritische Stellungnahme jedes Autors zu den Positionen der beiden anderen Autoren an. Zum Schluss folgt eine Replik jedes Einzelnen auf die Stellungnahmen. Dadurch fehlt allerdings ein klarer Duktus: Informationen werden auseinandergerissen, womit es schwerfällt, dem roten Faden zu folgen. Das Buch umfasst nicht ganz 400 Seiten.
Der Inhalt
Frau DROSSEL bespricht die „Lückentheorie“ (Gap Theory), die „Tag-Zeitalter-Theorie“ (Day-Age Theory) und die „schematische Sicht“ (Framework View), welche die Bedeutung der Schöpfungstage in der Bibel interpretieren (S. 31 ff.). Zur Frage nach der Abstammung des Menschen von anderen Primaten präsentiert sie einschlägige Belege wie Punktmutationen, Indels (S. 271), Multiplikation von Genen und anderen DNA-Abschnitten, Unterschiede im Y-Chromosom sowie De-novo-Gene (S. 272).
JUNKER deutet Konvergenzen, das heißt biologische Merkmale, die aus evolutiver Sicht in verschiedenen Entwicklungslinien unabhängig voneinander entstanden sind, als Ausdruck eines schöpferischen Baukastens (S. 148). Intelligent Design (ID) möchte dieser mit vermeintlichen Indizien, etwa mit nichtreduzierbarer Komplexität, spielerischer Komplexität und Plastizität, untermauert sehen (S. 151). Ferner behauptet er, der Design-Ansatz sei keine Konkurrenz zur Naturwissenschaft, sondern stehe lediglich in Konkurrenz zum Naturalismus (S. 306).
SCHERER stellt die reproduzierbare empirisch-experimentelle Naturwissenschaft der angeblich nicht falsifizierbaren empirisch-historischen Naturwissenschaft gegenüber. Angesprochen werden Homologien und Konvergenzen, Mikro- und Makroevolution. Des Weiteren widmet er sich theologischen Themen wie etwa der Teleologie in der Natur, der Frage der Theodizee und dem Sündenfall.
Im Folgenden werden einige Themen vorgestellt und kritisch besprochen.
Die Diskussion über das Alter der Erde
JUNKER äußert Zweifel an den etablierten Datierungsmethoden, die ein Erdenalter von rund 4,5 Milliarden Jahren liefern. Es überrascht nicht, dass er das Alter der Erde für „wissenschaftlich nicht endgültig beantwortet“ hält (S. 289). Er verweist auf Beobachtungen, die vermeintlich nicht zu den etablierten Altersdaten passen (S. 302 f.).
SCHERER hält zwar das hohe Alter der Erde und des Universums als „derzeit alternativlos“ (vgl. S. 310). Gleichwohl hält er „eine sachlich begründete Skepsis gegenüber herrschenden naturwissenschaftlichen Paradigmen“ für einen „zentrale[n] Bestandteil des wissenschaftlichen Forschungs- und Erkenntnisprozesses“ (S. 309). So erwähnt er „Funde von menschlichen Werkzeugen in bis zu 50 Mio. Jahre alten geologischen Formationen“, wobei er sich auf ein Buch von Michael BRANDT bezieht (vgl. S. 317). Darin gibt der Autor vor, nachgewiesen zu haben, dass es sich bei den uralten Steinen, deren Formen an primitive Steinartefakte erinnern, um Menschenwerk handele.
SCHERER lässt sein Urteil über den Artefaktcharakter der Steine offen. Er deutet nur an, die Funde würden eine „ziemlich ernsthafte Anfrage an gegenwärtige Theorien zur Evolution des Menschen“ richten (S. 317), sollten es Artefakte sein. Allerdings prüft er nicht sorgfältig den wissenschaftlichen Stellenwert von BRANDTs Interpretation, womit er eine Skepsis an den geologischen Zeiträumen nährt, die sachlich unbegründet ist.
Das Problem: Die Gründe, weshalb BRANDTs Deutung aus Expertensicht unhaltbar ist, sind lange bekannt (s. etwa ADRIAN 1948; NEUKAMM 2021). SCHERER lässt sie unerwähnt. Über diesen Sachverhalt muss er sich im Buch von DROSSEL aufklären lassen, ebenso über die kritische Literatur.
Irritierend ist, dass SCHERER anmerkt, es sei „wenig überraschend, dass die Einordnung solcher Funde als echte Artefakte in solchen Schichten bezweifelt wird“ (S. 317), ohne die Gründe zu nennen. Den Kontext, in den er seine Andeutung einbettet, wählt er so, dass der Eindruck entsteht, die Ablehnung erfolge wegen ihrer weltanschaulichen Brisanz. Das ist aber nicht der Fall. Selbst wenn einige der Steine Artefakte sein sollten, würde dies weder ihren menschlichen Ursprung noch eine junge Erde nahelegen. Ein solches Szenario ist hochgradig inkonsistent mit der übrigen Wissenschaft.
Die von JUNKER (S. 302 f.) angeführten Befunde gegen die radiometrischen Gesteinsdatierungen, wie etwa Gewebereste in Dinosaurierknochen, überzeugen Fachleute ebenso wenig. Es ist, wie DROSSEL anmerkt, „viel naheliegender, nach Mechanismen zu suchen, die solche Gewebe und Strukturproteine stabilisieren können“ (S. 91), als das gesamte in sich stimmige erdgeschichtliche Datierungssystem umzuwerfen. In der Tat haben die etablierten Naturwissenschaften für JUNKERs „Anomalien“ (S. 303) längst plausible Lösungen gefunden (vgl. z. B. SPRITKOPF 2020; NEUKAMM 2024),
„… ohne dass man sich mit der These einer kurzen Menschheitsgeschichte einen immensen Berg von neuen Problemen einhandeln muss“ (S. 341).
DROSSEL resümiert, dass es angesichts der erdrückenden Belege für ein nach Jahrmilliarden zu bemessenes Weltalter „irgendwann… schlichtweg unvernünftig“ wird, immerzu „an der akzeptierten Interpretation der Daten zu zweifeln“ (S. 340).
Widersprüchliche Stammbäume: Ein Argument gegen Evolution?
JUNKER vertritt das Ansinnen, wissenschaftlich am Fundament der evolutionären Stammesgeschichte zu rütteln. Er referiert Beispiele, wonach molekulare Daten zur Stammbaumrekonstruktion sicher geglaubte Verwandtschaftsverhältnisse infrage gestellt hätten, statt unklare Fälle zu lösen (vgl. S. 167). Stammbäume stünden regelmäßig im Widerspruch zueinander (Inkongruenz-Problem) und würden nach heutigem Wissen im Großen und Ganzen nicht gut zusammenpassen (S. 355).
Gewiss wird niemand bestreiten, dass die Rekonstruktion von Verwandtschaftsverhältnissen Schwierigkeiten bereitet. Doch warum sollte dies gegen die Historizität des Evolutionsgeschehens (bzw. gegen die Theorie gemeinsamer Abstammung) sprechen?
Was JUNKER nicht sagt: Das empirische Fundament, welches die evolutive Stammesgeschichte bezeugt, ist von den existierenden Inkongruenzen und Unsicherheiten über die exakte Stellung rezenter und fossiler Arten im Stammbaum gar nicht betroffen.
Die Historizität des Evolutionsgeschehens gründet wesentlich auf dem Befund, dass sich die Fossilien in eine Reihenfolge bringen lassen, in der sie mehr und mehr die Gestalt der heutigen Arten annehmen. Überwiegend korreliert die Reihenfolge gut mit der zeitlichen Abfolge in den Strata (BENTON 2003). Die Logik des Evolutionsbelegs liegt darin, dass dieses Muster den Erwartungen der Abstammungstheorie entspricht.
Dieser Beleg ist logisch unabhängig von der Frage, ob die Rekonstruktion der Verwandtschaftsverhältnisse durch Inkongruenzen erschwert wird, ob der Befund Diskontinuitäten anzeigt oder eine stark verzweigte Evolution nahelegt.
Betrachten wir zum Beispiel die vogelähnlichen Theropoden-Dinosaurier, so nimmt die Anzahl der Vogelmerkmale von Anchiornis über Archaeopteryx, Jeholornis, Confuciusornis und Ichthyornis zu den höheren Vögeln sukzessive zu (s. Abb. 1). Der Umstand, dass die genauen Verwandtschaftsverhältnisse der Taxa noch nicht geklärt sind, rüttelt nicht an der Tatsache, dass die sukzessive Annäherung der fossilen Arten an die rezente Vogelfauna ihre stammesgeschichtliche Entwicklung untermauert.

Abb. 1: Merkmalstabelle einiger Theropoden-Dinosaurier. (+) bedeutet: Merkmal vorhanden, (-): Merkmal fehlt, (±): Merkmal rudimentär vorhanden. Die orangefarbenen Felder zeigen den Besitz ursprünglicher Nichtvogel-Theropoden-Merkmale an, die grünen Felder die Präsenz fortschrittlicher Vogelmerkmale. Die Tatsache, dass im Lauf der Erdgeschichte immer vogelähnlichere Theropoden auftauchen, bildet ein wesentliches Fundament der Evolutionstheorie.
Keine eingeschachtelte Hierarchie von Merkmalen?
Setzt man voraus, dass die Theorie der gemeinsamen Abstammung richtig ist, ergibt sich logisch eine Hierarchie eingeschachtelter Merkmalsbeziehungen.
Zum Beispiel bilden alle Tiere mit einer Wirbelsäure (Wirbeltiere) auch eine Chorda (einen stabförmigen Stützapparat im Rücken) aus, aber nicht alle Tiere mit einer Chorda besitzen Wirbel. Alle Kiefermünder besitzen Wirbel, aber nicht alle Wirbeltiere sind Kiefermünder. Alle Fleischflosser sind Kiefermünder, aber nicht alle Kiefermünder sind Fleischflosser und so weiter. Diese eingeschachtelte Merkmalshierarchie erklärt sich schlüssig, wenn wir folgende Abstammungsverhältnisse annehmen: Die Fleischflosser entwickelten sich aus einem ursprünglichen Kiefermünder. Die Kiefermünder stammen von einem basalen Wirbeltier ab. Die Wirbeltiere wiederum evolvierten aus einem ursprünglichen Chordatier (s. Abb. 2).
Diese Erklärung stützt sich auf das Wissen um die genetische Variation von Merkmalen und um deren Vererbung. Eine schöpfungstheoretische Deutung ist dagegen nur mithilfe der willkürlichen Zusatzannahme zu haben, dass sich der Schöpfer dazu entschloss, die Arten über die „Grundtyp“-Ebene hinaus einander abgestuft ähnlich zu erschaffen. Eine vernünftige, wissenschaftliche Erklärung ist das freilich nicht.

Abb. 2: Vereinfachtes Verzweigungsschema (Kladogramm, Verwandtschaftsbaum) der Chordatiere bis zu den Fleischflossern, zu denen auch die Landwirbeltiere zählen. Es lässt sich anhand der eingeschachtelten Hierarchie von Merkmalsbeziehungen erstellen. Entsprechend dieser Hierarchie ist die Gruppe der Fleischflosser tief ins Taxon der Chordatiere eingeschachtelt. Das Kladogramm lässt sich in einen Stammbaum übersetzen, der eine Zeitachse enthält und die Arten nach dem Grad der Übereinstimmungen einordnet (Dendrogramm).
JUNKER bestreitet nun die Aussage der Mitautorin Barbara DROSSEL, dass der Vergleich der DNA verschiedener Arten zu einer eingeschachtelten Hierarchie von Ähnlichkeiten führe, die „zur Annahme eines Stammbaums passe und keine Anhaltspunkte für evolutionär voneinander getrennte Grundtypen liefere“ (S. 354):
„Tatsächlich sind… die Ähnlichkeitsbeziehungen verbreitet netzförmig und nicht eingeschachtelt. Diese Situation hat sich mit zunehmender Kenntnis verschärft. Beispielsweise ist die Situation bei der artenreichsten Pflanzengruppe, den bedecktsamigen Blütenpflanzen (Angiospermen), äußerst verwickelt: Dendrogramme (Ähnlichkeitsbäume) verschiedener Gene sind oft deutlich verschieden und das ganze System stellt sich überhaupt nicht eingeschachtelt dar – entgegen evolutionstheoretischen Vorhersagen.“ (S. 354)
Was JUNKER nicht sagt:
(1) Selbst bei den inkongruentesten Bäumen mit vielen „netzförmig“ verteilten Merkmalen ist die große Mehrheit der Merkmalsbeziehungen klar hierarchisch verschachtelt (s. THEOBALD 2013). Das gilt auch über vermeintliche „Grundtyp-Grenzen“ hinweg.
(2) „Netzförmige“ Merkmalsverteilungen sind auch innerhalb der sogenannten „Grundtypen“ verbreitet.
Ein Beispiel zu Punkt (1): Der Verwandtschaftsbaum der Pan-Aves umfasst neben den modernen Vögeln und „Urvögeln“ wie Archaeopteryx auch Nichtvogel-Dinosaurier, Flugsaurier und alle „Reptilien“, die näher mit den Vögeln verwandt sind als mit den Krokodilen. Dem Kreationismus zufolge müssten allein die rezenten Vögel bis zu 47 verschiedenen Grundtypen angehören, die sich wiederum wesensmäßig von den Dinosaurier- bzw. Reptilien-Grundtypen unterscheiden.
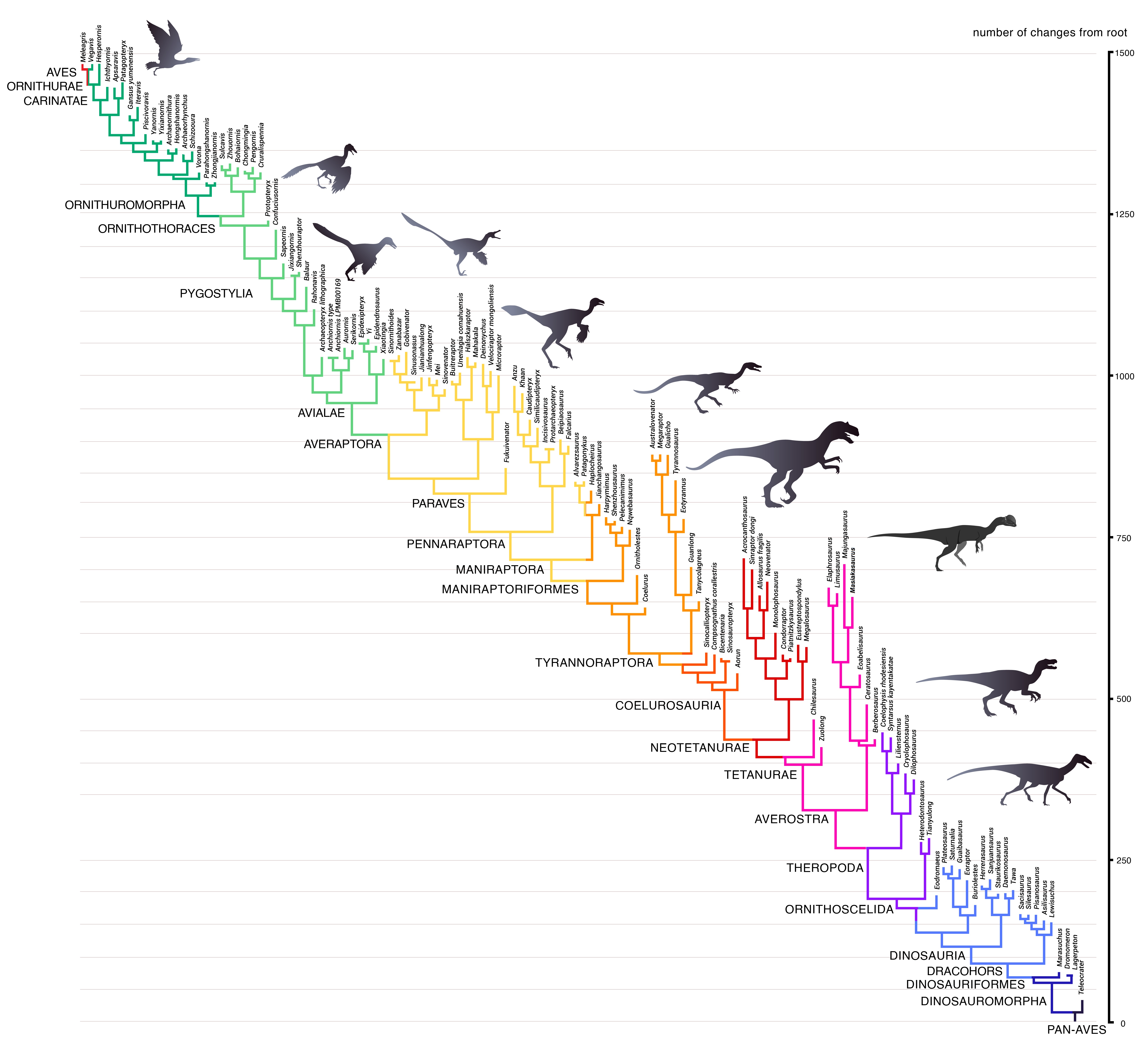{kind=link}
Nach CAU (2018) zeichnen sich die höheren Vögel gegenüber der Stammart an der Basis des Baums durch etwa 1500 morphologische Veränderungen (bzw. abgeleitete Merkmale) aus. Trotz Konvergenzen zeigt sich eine klare Merkmalshierarchie, die sich von den modernen Vögeln über die Nichtvogel-Dinosaurier bis zu den ursprünglichen Avemetatarsalia und darüber hinaus erstreckt. An keiner Stelle zeichnen sich Brüche ab, anhand derer sich „Grundtypen“ voneinander unterscheiden ließen.
An folgender Tatsache kommt JUNKER nicht vorbei:
„Im Allgemeinen können Stammbäume sehr inkongruent sein und dennoch in einem extrem hohen Grad an statistischer Signifikanz übereinstimmen… Der verblüffende Grad an Übereinstimmung selbst zwischen den inkongruentesten Stammbäumen, die wir aus der Literatur kennen, wird selten gewürdigt, hauptsächlich, weil die meisten Menschen (darunter viele Biologen) sich der betreffenden Mathematik nicht bewusst sind.“ (THEOBALD 2013; Dt. MN)
Selektives Zitieren verfälscht das Bild
Ein großes Problem ist, dass JUNKER seine Beispiele sehr selektiv wählt. So arbeitet er sich mit Vorliebe an Fällen ab, die ein völlig konfuses, uneinheitliches Bild zu vermitteln scheinen. Die große Mehrheit konsistenter Ergebnisse lässt er unberücksichtigt. In Wahrheit zeigen statistische Untersuchungen, dass trotz der erwähnten Probleme
„… die Daten der Molekularbiologie die vergleichende Morphologie in praktisch allen gut abgesicherten Fällen bestätigen…“ (MAIER 1994, S. 118)
Auch neuere Untersuchungen weisen darauf hin, „dass sowohl molekulare als auch morphologische Bäume im Allgemeinen brauchbare Annäherungen an eine gemeinsame zugrundeliegende stammesgeschichtliche Entwicklung liefern“ (PISANI et al. 2007, S. 269; Dt. MN).
PRASAD et al. (2008) untersuchten die verwandtschaftlichen Beziehungen zwischen 44 Säugetierarten anhand umfangreicher Sequenzdaten, die aktuelle molekulare phylogenetische Analysen weitestgehend bestätigten. Die Tatsache, dass unterschiedliche phylogenetische Datensätze immer wieder zu vergleichbaren Ergebnissen führen, die mit der Gesamtheit des Hintergrundwissens übereinstimmen, belegt, dass die Evolutionstheorie die Wirklichkeit adäquat beschreibt:
„Würden wir nämlich kein realhistorisches Ereignis rekonstruieren, hätten sich sicher viele Inkonsistenzen und Anomalien ergeben, die die Rekonstruktionsversuche schon im Ansatz hätten scheitern lassen… Die Theorie der Historizität eines Evolutionsgeschehens ist also so wohlbestätigt, dass wir es als Faktum bezeichnen können.“
MAHNER 1986, S. 40)
Mechanismen, die Inkongruenzen erzeugen
JUNKER bemerkt, diverse Teilgebiete der Biologie (Biogeografie) lieferten „kein durchweg einheitliches Bild“ (S. 304). Gerade er sollte wissen, dass biologische und insbesondere biohistorische Prozesse chaotisch sind, sodass immer mit einigen Unstimmigkeiten zu rechnen ist. Das gilt natürlich auch für das Inkongruenz-Problem. Die Gründe für die verschiedenen Disharmonien sind vielfältig und schon lange bekannt.
Gene müssen sich in verschiedenen Abstammungslinien nicht immer mit gleicher Geschwindigkeit entwickeln. Gelegentlich können Moleküle durch Selektion oder Zufall konvergieren. Immer wieder kommt es zu horizontalem Gentransfer. Auch Populationsgenetische Effekte wie unvollständige Liniensortierung (Incomplete lineage sorting) und Artbildung durch Hybridisierung erzeugen Inkongruenzen. Signalerosion, Genduplikationen und verborgene Paralogien verwischen die Phylogenien zusätzlich. Einen Überblick über die bekannten Konfliktquellen geben MAIER (1994); ISAAK (2007); COHEN (2018); SCHREMPF & SZÖLLÖSI (2020); HILL & ROCH (2022); STEENWYK et al. (2023).
Die Analyse morphologischer Strukturen kann zudem verrauschte oder konvergente Merkmale enthalten. Die Ursachen sind gleiche Selektionsfaktoren, Parallelismen und binäre Merkmalsausprägungen durch veränderte Umwelteinflüsse (Plastizität). Einige Dissonanzen beruhen auf der Strukturarmut mancher Taxa (SUDHAUS 2006). Da molekulare im Vergleich zu morphologischen Datensätzen ein reichhaltigeres Reservoir an phylogenetischer Information beinhalten, wäre es zudem seltsam, wenn morphologische und molekulare Daten immer die gleichen Stammbäume liefern würden.
Was JUNKER nicht sagt: Eine Fülle an Ursachen und Mechanismen lässt Inkongruenzen und Unstimmigkeiten aus evolutionärer Sicht geradezu erwarten. Wird dieser Aspekt nicht gewürdigt, ist eine kompetente Auseinandersetzung mit der Evolutionstheorie unmöglich.
Unstimmigkeiten entstehen auch aufgrund methodischer und interpretierender Fehler (SANDERSON & SHAFFER 2002). Die phylogenetische Analyse ist ein äußerst komplexes Thema; selbst von Experten kann man nicht erwarten, dass sie immer richtigliegen. Die Herausforderung besteht darin herauszufinden, welche Konfliktverursacher im Spiel sind, und die Modelle entsprechend anzupassen.
Eine Vielzahl an Unstimmigkeiten lässt sich beheben, indem man die Analyse auf so viele Gene wie möglich stützt (ROKAS et al. 2003) oder morphologische Datensätze mit molekularen Sequenzdaten kombiniert (s. etwa HERNÁNDEZ-LARA 2018; VAN DER WAL 2019; KEATING et al. 2023). Vor allem genomweite Datensätze können Genbaumkonflikte aufdecken und phylogenetische Beziehungen lösen, deren Klärung traditionell problembehaftet schien (SINGHAL et al. 2021).
TAUTZ (2006) betrachtet die Konflikte zwischen Stammbäumen, die aufgrund verschiedener Datensätze erstellt wurden, sogar explizit als wertvoll. Sie helfen nämlich dabei, Abstammungshypothesen zu testen und zu verfeinern – und vor allem die Mechanismen aufzudecken, die solche Inkongruenzen verursachen!
So kann sich unter Hinzuziehung entwicklungsbiologischen Zusatzwissens ein als homolog interpretiertes Merkmal auch als Parallelbildung herausstellen und umgekehrt. Dies ist ein klares Indiz dafür, dass prüfbare Verwandtschaftshypothesen erstellt werden können, die sich unter Einbeziehung des gesamten biologischen Hintergrundwissens bewähren müssen. Das schrittweise Herantasten an die Fakten durch wechselseitiges Klassifizieren und Theoretisieren ist kein Problemfall, sondern allgemein „ein Erkennungszeichen von Wissenschaft“ (MAHNER & BUNGE 2000, S. 246).
Fossilbefund, Stammbäume und „molekulare Uhren“
JUNKER behauptet, dass die molekularen Daten, anhand derer sich der Ursprung bestimmter Organismengruppen datieren lässt, mit dem Fossilienbericht oft nicht zusammenpassen. „Congruence between dates from molecular phylogenies and the fossil record“, so zitiert er einen Forscher zum Ursprung der Vögel, „is a rare thing“ (S. 167). Die Übergangsformen am Beginn der Radiation scheinen systematisch zu fehlen, so auch bei den Blütenpflanzen in der Kreidezeit. Generell wurde die
„… evolutionstheoretisch naheliegende Erwartung einer Übereinstimmung verschiedener Datensätze… vielfach enttäuscht, ohne dass dies zu kritischen Rückfragen an den evolutionären Ansatz geführt hätte.“ (S. 168)
Diese „kritischen Rückfragen“ sind für JUNKER gleichbedeutend mit Fundamentalkritik an der Evolutionstheorie. Freilich läuft diese Form der ideologisch motivierten „Rückfrage“ aus den genannten Gründen ins Leere.
Was JUNKER nicht sagt: Die Stammbäume korrelieren im Allgemeinen recht gut mit dem Fossilbefund (s. NORELL & NOVACEK 1992; BENTON 2003; WILLS et al. 2008; O'CONNOR & WILLS 2016). Man darf bei chaotischen Prozessen nur nicht erwarten, dass das immer der Fall ist.
Die Muster der Fossilienvielfalt können verzerrt sein, etwa durch unterschiedliche Erhaltungspotenziale von Organismen in verschiedener Umgebung, geologische Verzerrungen und heterogene Probenahme (FLANNERY-SUTHERLAND et al. 2022). Derlei Verzerrungen wirken sich aber meist nur dann aus, wenn man einzelne Arten an bestimmten Fundorten in feiner stratigrafischen Unterteilung betrachtet (BENTON 2003, S. 85).
Richtig ist, dass das Ergebnis der molekularen Datierung evolutiver Verzweigungen das Alter der entsprechenden Fossilien oft deutlich übersteigt. So legen phylogenetische Daten nahe, dass die Radiation der Vögel in der Kreidezeit stattfand; gemessen am Fossilbefund ereignete sich die Hauptradiation aber erst nach der Kreidezeit (THOMAS 2015). Der Ursprung der Blütenpflanzen wiederum wurde auf eine Zeit von vor 255 bis 206 Mio. Jahren berechnet (BARBA-MONTOYA et al. 2018), wogegen die ersten fossilen Vertreter der Blütenpflanzen mit 135 Mio. Jahren sehr viel jünger sind.
Der Grund: Die basalen Vertreter einer Gruppe waren häufig nicht artenreich und nicht verbreitet genug, um fossil belegt zu sein. Daher unterschätzt der Fossilnachweis regelmäßig das Alter der Divergenz (THOMAS 2015). Zudem weisen die Fossilien auf der Stammlinie einer Kronengruppe noch nicht alle definierenden Merkmale dieser Gruppe auf, sodass eine Zuordnung oft schwierig ist (SCHNECKENBURGER 2014). Bezogen auf die Blütenpflanzen findet man in der frühen Kreide eine Fülle von Fossilien, die eine Zunahme struktureller Komplexität belegen. Es handelt sich um Formen, die noch zu keiner rezenten Familie gehören. Älterer Pollen mit einem den Blütenpflanzen ähnlichen Bau ist schon länger bekannt (CORNET & HABIB 1992; TAYLOR et al. 2006).
Was Junker nicht sagt:
(1) Bei den Vögeln haben jüngste Funde, wie der kürzlich beschriebene Asteriornis, die Diskrepanz zwischen molekularen Daten und Fossilbefund weitestgehend aufgehoben (FIELD 2020, s. Abb. 3).
(2) Inzwischen wird ein vorkreidezeitlicher Ursprung der Blütenpflanzen nicht nur durch molekulare Uhren, sondern auch durch die Analyse von Fossilienfunden gestützt. Die Ergebnisse „stimmen bemerkenswert gut mit den jüngsten Schätzungen zur molekularen Uhr für das Kronenalter der Angiospermen überein“ (SILVESTRO et al. 2021, S. 454; Dt. MN).

Bild: Schoepfung-Evolution Abb. 3
Abb. 3: Heute unterstützen sowohl der Fossilbefund als auch die zeitlich kalibrierten molekularen Stammbäume das gleiche Szenario: Die neuzeitlichen Vögel entstanden während der späten Kreidezeit, allerdings nicht erst an deren Ende. Dieser Stammbaum zeigt, dass sich die Hauptkladen der neuzeitlichen Vögel bereits vor rund 100 Millionen Jahren begannen, auseinanderzuentwickeln. Am Ende der Kreidezeit hatten sich schon diverse Abstammungslinien etabliert. Unbestritten ist freilich, dass der größte Teil erst nach der Kreidezeit entstand. Aus FIELD et al. (2020).
Solche Beispiele zeigen, wie die Forschung paläontologische und molekularphylogenetische Diskrepanzen zur Evolutionsgeschichte sukzessive aufhebt. Es gibt keine belastbaren Hinweise auf eine Entkopplung von molekularer und morphologischer Evolution (OMLAND 1997).
Konvergenz: Design-Indiz oder Ausdruck kanalisierender Evolution?
Zum Thema „Konvergenz“ schreibt JUNKER:
„Konvergenz muss häufig auch bei komplexen Konstruktionen oder sogar bei Schlüsselmerkmalen angenommen werden. Letztere sind Merkmale, die definierend für größere Organismengruppen sind, wie z. B. die drei Gehörknöchelchen der Säugetiere. Mittlerweile ist klar: Ein verbreitetes Vorkommen von Konvergenzen ist normal.“ (S. 148)
Andererseits könnten Konvergenzen als Wiederverwendung bestimmter Bauplanmodule oder Konstruktionsideen interpretiert werden. Das sei sehr gut verständlich im Rahmen eines Schöpfungskonzepts. Ein „genialer Schöpfer“ könne eine Bauplanidee
„… beliebig oft in verschiedensten Zusammenhängen verwenden, wo immer er möchte [sic!] oder es für zweckmäßig hält. Mehrfachanwendungen und daraus folgend eine Art ‚Baukastensystem‘ sind typische Kennzeichen ausgeklügelter Planung, also Schöpfungsindizien.“ (S. 148)
Das Problem: Da wir weder über den Schöpfer noch über dessen Handlungsmodus auch nur das Geringste wissen, beruhen derlei „Schöpfungsindizien“ auf willkürlichen Ad-hoc-Annahmen. Die Evolutionstheorie liefert die schlüssigere Antwort, da ähnliche Selektionsdrücke und die Kanalisierung der Evolution entlang gleicher Entwicklungsbahnen bekanntermaßen Konvergenzen (Parallelismen) erzeugen (MCGHEE 2011, S. 7).
Kanalisierung bedeutet, dass die historisch gewachsenen strukturellen Zusammenhänge in einem Merkmalskomplex oder Bauplan nur noch bestimmte Modifikationen zulassen. Die evolutionär getroffenen Vorentscheidungen üben „Zwänge“ auf die weitere evolutionäre Entwicklung aus, indem sie die möglichen Entwicklungswege einschränken. Dies kann zweierlei Auswirkungen haben: Auf der einen Seite können
„… bestimmte Strukturen nie entstehen…, weil es objektiv gewisse innere (biochemische, physikalische oder auf der Konstruktion beruhende Zwänge) gibt, die die Evolution nicht überwinden kann.“ (ZRZAVÝ et al. 2013, S. 270)
Auf der anderen Seite kann genetische Beschränkung zur Konvergenzbildung beitragen. Die wiederholte, unabhängige Rekrutierung eines bestimmten genetischen Moduls zur Kontrolle eines Merkmals kann die Tatsache widerspiegeln, dass die zugrundeliegenden Entwicklungsprozesse bestimmten Zwängen unterliegen (HODIN 2000).
Ein gut untersuchtes Beispiel betrifft die genetische Kontrolle der Blattmorphologie, eines der variabelsten Merkmale bei Pflanzen. Die Bildung von zusammengesetzten Blättern hat sich allein bei den Blütenpflanzen dutzende Male unabhängig von ihren einfachblättrigen Vorfahren entwickelt.
Entwicklungsgenetisch lässt sich dies gut erklären: Was fast alle diese zusammengesetzten Blätter gemeinsam haben, ist die Expression bestimmter KNOX-Gene, die mit dem teilungsfähigen Bildungsgewebe der Pflanzen assoziiert sind.
„Es hat den Anschein, dass eine tief konservierte Rolle bei der Aufrechterhaltung der Identität des teilungsfähigen Bildungsgewebes den KNOX-Signalweg dazu veranlasst hat, viele Male unabhängig rekrutiert zu werden… Eine besonders elegante aktuelle Studie von Hay und Tsiantis (2006) hat entscheidende Beweise dafür geliefert, dass der KNOX-Signalweg unabhängig rekrutiert wurde, um die Unbestimmtheit in Blättern zu fördern, und deutet darüber hinaus auf ein hohes Maß an Labilität in den Regulationswegen hin, die KNOX-Gene in einem typischen einfachen Blatt unterdrücken… Dieses und andere Beispiele… scheinen in die Klasse der Parallelität zu passen…“ (KALISZ & KRAMER 2008, S. 174; Dt. MN).
Ist Junk-DNA funktional?
Das „Junk-DNA-Argument“ (also die Ansicht, dass der größte Teil unseres Genoms nicht funktional ist, vgl. S. 163) kontert JUNKER mit einem Verweis auf das ENCODE-Projekt. Dieses habe aufgedeckt, dass große Teile des Erbguts transkribiert (in mRNA umgeschrieben) würden, mithin funktional seien und nicht als evolutiver Müll interpretiert werden könnten (vgl. S. 168). Augenscheinlich vermengt und verwechselt JUNKER „Transkription“ mit „Funktionalität“ – eine schwere Fehleinschätzung, wie die Autoren des ENCODE-Projekts selbst betont haben:
„A priori, we should not expect that transcriptome to consist exclusively of functional RNAs. Zero tolerance for errant transcripts would come at high cost in the proofreading machinery needed to perfectly gate RNA polymerase and splicing activities, or to instantly eliminate spurious transcripts. In general, sequences encoding RNAs transcribed by noisy transcriptional machinery are expected to be less constrained, which is consistent with data shown here for very low abundance RNA.“ (KELLIS et al. 2014; Hervorhebung hinzugefügt.)
LYNCH & MARINOV (2015) haben empirisch bestätigt, dass die bioenergetischen Kosten zur Entfernung falscher Transkripte unterhalb eines bestimmten Levels zu niedrig sind, um effektiv dagegen zu selektieren. Die Kosten sind aber meist hoch genug, um deren Translation zu unterdrücken.
Entscheidend ist nun, dass die von ENCODE selbst genannten Indizien für nichtfunktionale RNA (less constrained und very low abundance) nicht nur einige wenige Transkripte betreffen, sondern die überwältigende Mehrheit des Transkriptoms!
Was JUNKER nicht sagt: 92 % des Genoms sind „unconstrained“ (RANDS et al. 2014). Mutationen können sich im Lauf der Zeit ansammeln, ohne dass Teile der Sequenz durch reinigende Selektion konserviert bleiben. Des Weiteren weisen 90 % aller RNA-Transkripte eine Konzentration zwischen 0,1 und 1 Molekülen pro Zelle auf (Abb. 4). Beides sind nach ENCODE starke Indizien für Nichtfunktionalität.

Abb. 4: Abdeckung des menschlichen Genoms durch die ENCODE-Daten. Der Anteil des menschlichen Genoms, der durch ENCODE-Elemente in mindestens einer Zelllinie oder einem Gewebe für jeden Test abgedeckt wird, ist in einem Balkendiagramm dargestellt. Alle Prozentsätze werden für das gesamte Genom berechnet, einschließlich des Teils, der mit kurzen Reads nicht eindeutig zugeordnet werden kann und daher für die hier vorgestellte Analyse unsichtbar ist. Quelle: KELLIS et al. (2014, S. 6133).
Nun könnte man spekulieren, dass solche Low-level-Transkripte in einem bestimmten Zelltyp in einer höheren Konzentration vorliegen und somit doch funktional sein könnten. Allerdings wurde bereits die zelllinienspezifische Expression von Retrotransposons (LINEs, SINEs und LTRs) untersucht (DJEBALI et al. 2012), die zusammen fast 50 % des Genoms ausmachen. Das Ergebnis stützt die Spekulation nicht (s. Abb. 5).

Abb. 5: Zelllinienspezifische Expression von Repeat-Elementen im menschlichen Transkriptom. Die Heatmaps erfassen alle LINEs, SINEs und LTR-Repeats, die in mindestens einem Zelltyp oder Zellkompartiment in einer Konzentration von ≥ 1 pro eine Million RNA-Transkripte exprimiert werden. Jede Spalte steht für ein anderes Transkript (Repeat-Kopie). Jede Zeile wiederum steht für eine andere Zelllinie bzw. ein anderes Zellkompartiment. Quelle: DJEBALI et al. (2012), Supplementary Figure S20.
Nehmen wir zu JUNKERs Gunsten an, ein Expressionslevel von 1 pro eine Million RNA-Transkripte in einem beliebigen (!) Zelltyp würde bereits für eine Funktion reichen. Nehmen wir ferner an: Jedes gerade noch aufgelöste Bild-Pixel in der Originalarbeit (Abb. 5) repräsentiere ein anderes Repeat-Element. Dann kämen wir durch Auszählung der horizontalen Pixel auf grob 5.000 funktionale Repeat-Elemente. Im Verhältnis zu den 1,7 Millionen LINEs, SINEs und LTRs im Säugetier-Genom wären damit weniger als 0,3 % der Repeat-Elemente funktional!
Gewiss könnte man weiter spekulieren, dass für 99,7 % (!) der Retrotransposons irgendwann noch eine Funktion gefunden werde. Ein solches „Argument“ ist aber Lichtjahre von der starken Behauptung entfernt, dass „große Teile des Erbguts“ derzeit „nicht als evolutiver Müll interpretiert werden können“. Der Schluss auf die Funktionslosigkeit großer DNA-Bereiche beruht nach heutiger Faktenlage auf positivem Wissen.
Teleologie: Evolvierten Vogelflügel zum Fliegen?
SCHERER behauptet, das Profil und die Geometrie des Storchenflügels seien teleologisch auf einen Zweck und ein Ziel hin ausgerichtet. Der Vogelflügel sei so beschaffen, dass die Luftströmung einen Auftrieb erzeuge. Das Ziel bestehe in der Befähigung des Vogels zum Fliegen, der Zweck wiederum in der Futtersuche, im Schutz vor Fressfeinden, im Vogelzug etc. (S. 213).
Was SCHERER nicht sagt: Die nur wenig an einen Flug angepassten Flügelprofile und Tragflächen diverser Dinosaurier (etwa Yi, Anchiornis) sowie die unterschiedlichsten Flugarten, mit denen sie experimentierten, passen ins Bild eines chaotischen, nicht intendierten Naturprozesses. Auch die Tatsache, dass die rudimentären Flugversuche durchweg in Sackgassen endeten, spricht gegen einen teleologischen Ursprung.
Ein Großteil der Merkmale, die Vögeln das Fliegen ermöglichen, existierte bereits in ursprünglichen Coelurosauriern. Sie entstanden weder simultan, noch waren sie auf einen Flug hin „ausgerichtet“. Sie bildeten sich sukzessive und isoliert voneinander und standen im Dienste unterschiedlichster Funktionen. Selbst so charakteristische Vogelmerkmale wie Konturfedern und Flügel waren primär nicht von der Selektion auf aerodynamische Eigenschaften geprägt. Sie erfüllten andere Aufgaben wie Wärmeisolation, Balz, Imponier- oder Brutverhalten (BRUSATTE 2017, S. 793).
Erst später schufen die Merkmale in der Summe ganz beiläufig (zufällig) die Voraussetzungen für einen rudimentären Flug. Sie brauchten nur mit einem Sprungverhalten kombiniert und auf die neue Funktion hin optimiert zu werden (s. FOTH 2008).
Nichtreduzierbare Komplexität: Ein Design-Indiz?
JUNKER diskutiert die sogenannte nichtreduzierbare Komplexität als eines der Hauptargumente für Intelligent Design (S. 151). Was versteht man darunter?
„Ein nichtreduzierbar komplexes System ist per Definition eine komplizierte Struktur, deren Bausteine derart unabdingbar für die Funktion des Ganzen sind, dass jedwede Änderung, geschweige denn ein Verlust, alles Übrige völlig funktionsunfähig macht.“ (ZRZAVÝ et al. 2013, S. 472)
Da sich nichtreduzierbar komplexe Systeme an keiner Stelle vereinfachen lassen, ohne dass die biologische Funktion wegbricht, sei es unmöglich, sie in hinreichend kleinen, von der Selektion belohnten Schritten aufzubauen. Die Evolution müsse alle Bauelemente gleichzeitig bereitstellen und passend verschalten. Zudem müsse sie alle Teile hochgradig speziell feinjustieren, um die Funktion zu gewährleisten.
Schon bei einfachen Systemen soll dies vielfache Abstimmungen gleichzeitig erfordern. Daher, so meint JUNKER, sei es „sehr unwahrscheinlich, dass eine funktional-komplexe Struktur überhaupt ein einziges Mal entsteht (vgl. Design-Indizien)“ (S. 148). Nur Intelligenz und vorausschauendes Planen könnten das leisten. Insgesamt hält JUNKER „eine Makroevolution – gemessen an den insgesamt heute vorliegenden naturwissenschaftlichen Daten“ für „ausgesprochen unglaubwürdig“ (S. 289).
Was JUNKER nicht sagt: Nichtreduzierbar komplexe Systeme können durchaus graduell entstehen, etwa indem einer Vorläuferstruktur sukzessive Elemente hinzugefügt werden, welche die ursprüngliche Funktion verbessern (ZRZAVÝ et al. 2013, S. 288). Ab einem bestimmten Punkt kann der Merkmalskomplex neben der ursprünglichen Funktion unvermittelt eine neue, abgeleitete Funktion ausüben. Dies erörtern z. B. VOLLMER (1986, S. 25); MÜLLER (1990); DRAPER (2002); ZRZAVÝ et al. (2013).
ZRZAVÝ et al. (2013, Kap. 4.18) unterfüttern dieses Szenario mit Beispielen aus der Biologie. Dabei erfüllen einige Merkmale neben ihrer ursprünglichen Aufgabe eine Brückenfunktion. Wird die ursprüngliche Aufgabe in einem anderen Kontext bedeutungslos, können ursprüngliche Elemente verloren gehen, sodass im Ergebnis die verbleibenden Elemente unentbehrlich für das Gesamtsystem werden:
„Das, was am Anfang eine Luxusverbesserung war, wird zur Notwendigkeit. So ist ein nichtreduzierbares komplexes System entstanden, aber nicht durch das Zusammenfügen von ursprünglich unabhängigen Bestandteilen.“ (ZRZAVÝ et al. 2013, S. 288)
Ein weiterer Punkt, den JUNKER ignoriert: Ein hoher Grad an nichtreduzierbarer Komplexität in einem hochspezialisierten System sagt nichts darüber, wie komplex das Arrangement in einem noch wenig spezialisierten „Mini-System“ sein kann. Es zeigt sich immer wieder, dass die Teile der betreffenden Systeme nicht zwingend von Anfang an existieren und hochgradig speziell ausgeführt sein müssen (s. NEUKAMM 2022).
MILLER (1999) erörtert am Beispiel der Blutgerinnungskaskade, wie sie schrittweise entstanden sein kann, und führt JUNKERs Behauptung, solche Systeme seien „Hürden, die durch Mutation und Selektion nicht genommen werden können“ (S. 141), ad absurdum.
Aufschlussreich ist in diesem Zusammenhang eine bemerkenswerte Meinungsrevision: JUNKER & SCHERER (2006, S. 185) bezeichnen die Evolution der eukaryotischen Zelle, wie sie die Endosymbionten-Theorie beschreibt, noch als eine sehr unwahrscheinliche, „konzertierte Aktion“. Das System erscheine „irreduzibel komplex“ und wirke „als Design-Signal“. NEUKAMM & BEYER (2011) haben gezeigt, dass das nicht stimmt. Daraufhin räumten JUNKER & SCHERER (2013, S. 199) ein, dass die Datenlage „eine schrittweise Entwicklung zu Endosymbionten zunehmend plausibel“ mache. Wohlgemerkt, von einem „Design-Signal ist jetzt keine Rede mehr!
Selbst wenn JUNKER Recht hätte damit, dass die Evolution auf zwei, drei oder mehr „passende“ Mutationen „warten“ müsse (was nicht der Fall ist), würde das keineswegs bedeuten, dass derlei nur „sehr selten“ (S. 141) auftreten könne. Zum einen existieren schon heute empirische Befunde, die dem klar widersprechen (Weiteres dazu unten). Zum anderen:
Was JUNKER nicht sagt: Monte-Carlo-Simulationen legen nahe, dass unter Berücksichtigung kryptischer Variation gerade bei sich sexuell reproduzierenden Organismen die Chancen hoch sind, irreduzibel komplexe Systeme zu erzeugen – selbst wenn zwei, drei oder mehr „passende“ Mutationen dafür erforderlich wären (s. TROTTER et al. 2014).
Kryptische Variationen sind Mutationen in DNA-Regionen, die keinen Einfluss auf den Phänotyp haben, aber unter bestimmten Bedingungen einen solchen Effekt haben können. Dazu zählen z. B. Mutationen in Pseudogenen.
SCHERER und die Bakterienflagelle
Viele Bakterien besitzen eine „Rudergeißel“, ein Flagellum, mit dem sie sich fortbewegen. Dieses Flagellum ist nichtreduzierbar komplex: Nahezu alle Bestandteile (das heißt: mehrere Dutzend Proteine) werden für ihre Funktion benötigt. Darum ist das Flagellum eines der Standardbeispiele des intelligenten Designs.
SCHERER schreibt: Wenn es in Zukunft gelänge, die Entstehung biomolekularer Maschinen wie des bakteriellen Flagellums durch materielle Evolutionsmechanismen schlüssig zu erklären, wäre diese ID-Voraussage falsifiziert (vgl. S. 225). Doch entgegen SCHERER hat die Evolutionsbiologie längst begründete Vorstellungen darüber, wie Flagellen entstanden sind, auch wenn sie nicht alle Einzelschritte erklären kann.
Was SCHERER nicht sagt: Das heute favorisierte Modell zur Evolution von Bakterienflagellen (MATZKE 2006) verwickelte ihn bereits vor Jahren in ein Rückzugsgefecht (s. PEITZ 2013). SCHERER (2010) hält dieses Modell zwar für unzureichend. Es ist aber offenbar schon hinreichend genug, um ihn zur Revision seiner in JUNKER & SCHERER (2006, S. 162) vertretenen These zu zwingen, die Evolution der Flagelle sei transastronomisch unwahrscheinlich (Details in NEUKAMM et al. 2013).
SCHERER (2010) konzediert, dass anstatt der in JUNKER & SCHERER (2006, S. 162) noch postulierten 160 (!) nur noch etwa 10 Mutationen zwischen zwei positiv selektierbaren Vorstufen der Flagellen liegen sollen. Dies allein wäre schon ein erhebliches Zugeständnis. Doch auch die neu postulierte 10-fach-Mutation ist nur ein Asylum ignorantiae, ein „Zufluchtsort der Unwissenheit“.
Ferner bleibt die Frage, ob selbst eine „schlüssige“ materielle Erklärung – was SCHERER auch immer darunter verstehen mag – Intelligent Design tatsächlich falsifizieren würde. Dass dies nicht der Fall ist, lässt sich BEHEs eigenen Worten entnehmen:
„Und obwohl einige religiöse Denker sich ein aktives, ständiges Eingreifen in die Natur vorstellen, ist intelligentes Design durchaus mit der Ansicht vereinbar, dass das Universum ohne Überschreitung der Naturgesetze funktioniert, wobei das Design des Lebens vielleicht in seine ursprüngliche [!] Struktur gepackt wurde.“ (BEHE 2008, S. 166; Dt. MN)
Der Schluss auf Design als Schluss auf die beste Erklärung
Intelligent Design behauptet, bei nichtreduzierbar komplexen Systemen sei der Schluss auf eine intelligente Verursachung derzeit die beste Erklärung. Dieser sachlich begründete Anspruch sei eine direkte Aufforderung an Kosmologen und Evolutionsbiologen, einen überzeugenderen, ausschließlich auf materielle Prozesse gegründeten Entstehungsmechanismus vorzuschlagen (vgl. S. 222).
Intelligent Design wird hier jedoch ungerechtfertigterweise als Standard vorausgesetzt. Warum der Schluss auf „Design“ bei Naturgegenständen kein Schluss auf die beste Erklärung sein kann, erörtern z. B. MAHNER (2018) und NEUKAMM (2022).
DAWKINS' Wiesel-Programm: Kumulative Selektion und Teleologie
Der Begriff der „kumulativen Selektion“ wurde von Richard DAWKINS geprägt. Er meint damit den sich aufsummierenden Effekt vieler nacheinander passierender Mutationen unter einem richtenden Selektionsdruck.
SCHERER glaubt, dass die Darwin‘sche Evolution keine Strukturen erzeuge, die so aussehen, als seien sie durch Design entstanden. Zwar erkennt er kumulative Selektion „unter bestimmten Voraussetzungen“ an (vgl. S. 218 f.). Diese formuliert er allerdings so restriktiv, dass nur relativ banale Anpassungen darunterfallen.
SCHERER verweist auf DAWKINS' berühmtes „Wiesel-Programm“, welches er im dritten Kapitel seines Buchs „Der blinde Uhrmacher“ vorstellt. DAWKINS erläutert darin das Prinzip der Darwin‘schen Evolution mithilfe eines Computerprogramms. Am Anfang erzeugt das Programm eine Zufallsfolge aus 28 Buchstaben. Die Anzahl der möglichen Buchstaben-Kombinationen mit dieser „Satzlänge“ beträgt etwa 1040. Es erscheint demnach praktisch aussichtslos, durch „blindes“ Herumhämmern auf einer Schreibmaschine auch nur einen sinnvollen Satz aus 28 Buchstaben zu erzeugen.
Nun simuliert DAWKINS' Programm das Wirken natürlicher Selektion. Dazu gibt sein Programm einen Zielsatz vor. Steht ein Buchstabe zufällig an der richtigen Stelle, wird dieser „selektiv festgehalten“, während die übrigen erneut variieren. Anschließend werden wieder alle Buchstaben, die sich an den richtigen Stellen befinden, „fixiert“ und so fort. So zeigt DAWKINS, dass der Computer nach nur 43 Runden der Variation und Selektion aus dem Buchstabensalat den Zielsatz zu reproduzieren in der Lage war – ein Ergebnis, welches „allein durch Zufall“ nie hätte entstehen können.
SCHERER behauptet nun, DAWKINS habe sein Beispiel „wenig bedachtsam“ gewählt. Die Vorgabe eines Zielsatzes, an dem jede Zwischenstufe „gemessen“ werde, sei ausgerechnet das Paradebeispiel eines teleologischen Prozesses. Außerdem müsse das Computerprogramm (in der Biologie das evolvierende Lebewesen) irgendwo herkommen (vgl. S. 218).
Was SCHERER nicht sagt: DAWKINS' Programm gibt nur aus methodischen Gründen einen Zielsatz vor, nicht aus ontologischen: Wie sollen algorithmisch Selektionskriterien simuliert werden, wenn die „Passungen“ nicht im Programmcode hinterlegt sind?
In der Natur ist die Vorgabe von Zielen gänzlich unnötig, da in ihr erst durch die Interaktion des Organismus mit seiner Umgebung vorteilhafte „Ziele“ eruiert werden. Das bedeutet schlicht: Erweisen sich Mutationen als vorteilhaft, haben sie statistisch eine größere Chance, sich in Populationen durchzusetzen als neutrale oder schädliche Mutationen. Stellen wir fest, dass sie sich tatsächlich durchgesetzt haben, sind sie einer selektionstheoretischen Ex-post-facto-Erklärung zugänglich.
SCHERERs Kritik greift aus einem weiteren Grund daneben: Das Programm dient in diesem Szenario lediglich der Abbildung des Evolutionsprozesses, der selbst keinen teleologischen Aspekt beisteuert. Selbst wenn man argumentiert, dass das Programm einen Schöpfer benötige, läuft das in SCHERERs Analogie darauf hinaus, dass ein Schöpfer nur den Rahmen für eine natürlich evolvierende Welt bereitstellen müsste. Keinesfalls würde dies schöpferische Eingriffe erforderlich machen.
Im Übrigen ist die moderne Evolutionsbiologie längst über das alte Darwin‘sche Erklärungsparadigma hinausgelangt. Sie benötigt weder einseitig „selektionistische“ noch adaptationistische Erklärungen. Zahllose Merkmale in der belebten Natur sind nicht durch Selektion, sondern durch Drift in Populationen verankert worden. Solange die zufällig beibehaltenen Merkmale nicht unmittelbar tödlich für den Organismus sind, können sie zum Ausgangspunkt für neue „Evolutionsexperimente“ werden.
Das kreative Potenzial der Evolution
Im Hinblick auf die essentielle, aber seiner Meinung nach begrenzte Bedeutung der Gene für die Evolution und die damit einhergehenden Erklärungsdefizite der Synthetischen Theorie der Evolution hält es SCHERER für wahrscheinlich, dass die Diskussion auf einer anderen Ebene wiederaufleben werde (vgl. S. 214). Damit meint er die Diskussion über die Evolutionäre Entwicklungsbiologie, kurz: Evo-Devo (vgl. S. 392).
Bezeichnenderweise räumen JUNKER und SCHERER Evo-Devo einen weit niedrigeren Stellenwert ein als Fachleute. Zwar stimmen sie deren Aussagen über offene Detailfragen und die Probleme althergebrachter, selektionstheoretischer Erklärungen zu. „Knackigere“ Äußerungen wie die Folgenden dürften sie jedoch kaum unterschreiben:
„Tatsächlich räumen die neuen Fakten und Erkenntnisse aus der Entwicklungsbiologie und der Evo-Devo mit den kläglichen Überresten einer abgedroschenen, die Evolution leugnenden Rhetorik über die Bedeutung von Zwischenformen oder die Wahrscheinlichkeit, dass sich komplexe Strukturen entwickeln, auf. Wir haben inzwischen eine Vorstellung davon, wie sich von einer einzelnen Zelle aus die Komplexität eines ausgewachsenen Tieres entwickelt. Außerdem können wir mithilfe einer Reihe vollkommen neuer effizienter Methoden beobachten, wie sich durch Veränderungen in der Embryogenese die Komplexität und Vielfalt erhöhen. Die Entdeckung des uralten genetischen Werkzeugkastens ist ein eindeutiger Beleg dafür, dass Tiere wie Menschen von einem einfachen gemeinsamen Vorfahren abstammen, der modifiziert wurde.“ (CARROLL 2008, S. 18 f.)
Wenn man freilich so weit geht wie JUNKER, der jedwedes im Genom schlummernde Entwicklungspotenzial als „Mikroevolution“ und „programmierte Variation“[1] deutet (vgl. S. 106 f.); wenn man ferner behauptet, dass „echte“ Neuheiten „nicht auf bereits vorhandene, latente Anlagen zurückgeführt werden“ könnten (Anmerkung 73, S. 153), wird man immer die Prämisse bestätigt sehen, dass Evolution nichts „wirklich“ Neues hervorbringen könne.
Was JUNKER nicht sagt: Auch die weitreichendste evolutive Neuheit kann nur auf „bereits vorhandene, latente Anlagen zurückgeführt werden“ (s. JESSBERGER 1990). Den Grundstein legt zum Beispiel die Duplikation eines bereits vorhandenen Gens, welches dann solange abgewandelt wird, bis eine neue Funktion entsteht (Neofunktionalisierung).
Exemplarisch haben wir dies am Beispiel der Entstehung der Vögel aus ursprünglichen Dinosauriern erörtert – einer Evolution, die Kreationisten gemeinhin als Makroevolution auffassen (NEUKAMM & BEYER 2023, Abschn. C. 5 und C. 6).
Wissenschaftler haben heute gut begründete Vorstellungen darüber, wie die vogeltypischen Charakteristika entstanden sind. Deren Anlagen lassen sich teils bis zu den frühen Dinosauriern zurückverfolgen. Duplikation und Differenzierung „bereits vorhandener“ Gene spielten dabei ebenso eine Rolle wie Veränderungen der zeitlichen Koordination von Entwicklungsprozessen und andere genregulatorische Änderungen. Dabei vollzogen sich so wichtige Entwicklungsschritte wie die Verringerung der Körpergröße, die Zunahme des Stoffwechsels, die relative Zunahme des Hirnschädelvolumens, die Entstehung von Hornschnäbeln, Konturfedern und Brustbeinkamm, die Reduktion von Zähnen und Fingern, die Verschmelzung von Mittelhandknochen und vieles andere.
Entweder degradiert man nun die Entstehung der Vögel aus mesozoischen Dinosauriern zur Mikroevolution, was aus verschiedenen Gründen niemanden überzeugen würde. Oder man räumt ein, dass die Evolutionsbiologie gut begründete Vorstellungen darüber hat, wie sich makroevolutive Entstehungsprozesse aus vorhandenen, latenten Anlagen entfalten.
Ein weiterer Kritikpunkt ist, dass JUNKERs Konzept der „programmierten Variation“ eine nicht überprüfbare, metaphysische Ad-hoc-Annahme darstellt (s. Thomas JUNKER 2009). Sie ist nur dazu da, die Prämisse, Evolution könne nichts Neues hervorbringen, im Hinblick auf die Kenntnis unzähliger Variationsmechanismen zu retten, die das Potenzial dazu haben. Vor diesem Potenzial verschließt JUNKER konsequent die Augen: „Ein kreatives Potenzial von Evolution ist nicht erkennbar“, behauptet er holzschnittartig (S. 349).
Man ist versucht, ihm einen der vielen dokumentierten Fälle von Neofunktionalisierung entgegenzuhalten. Beispielsweise zeigen DE KRAKER & GERSHENZON (2011), dass durch zufällige Deletion eines für 120 Aminosäuren kodierenden Stücks DNA im Genom der Ackerschmalwand aus einem Protein ein neues Enzym mit komplett veränderter Architektur entstand. Der Clou: Dieses Enzym katalysiert zufällig die Bildung von Senfölglykosiden und schützt dadurch die Pflanze vor Raupenfraß.
Ohne Zweifel handelt es sich hier um eine nicht absehbare Neuerung mit hohem adaptivem Wert! JUNKER aber lässt solche Fälle nonchalant in der metaphysischen Schublade der „programmierten Variation“ verschwinden, wenngleich diese Deutung schon aufgrund des a priori unwahrscheinlichen Mutationsereignisses unplausibel ist.
Weniger leicht dürfte ihm dies im folgenden Fall gelingen: Im Lauf der Züchtung verschiedener Maissorten verschmolzen in einer Linie zwei Sequenzen des Mitochondrien-Genoms miteinander. Dabei entstand ein chimäres Protein, das einen nichtreduzierbar komplexen (!) Ionenkanal formt. Das Protein namens „T-urf13“ besitzt spezifische Bindestellen und Strukturelemente, die mit weiteren Einheiten das funktionelle System bilden. Bestimmte Moleküle öffnen oder schließen den Ionenkanal.
Der Clou: Die Sequenzen, aus denen T-urf13 entstand, enthielten davor niemals ein proteinkodierendes Leseraster – das Protein entstand de novo in sehr kurzer Zeit. Wir haben diese Evolution recht umfangreich dokumentiert (s. BEYER 2018; BEYER et al. 2022).
Ein anderer Fall, die Entstehung des „nichtreduzierbar komplexen“ Systems der eukaryotischen Zelle, wie sie die Endosymbionten-Theorie beschreibt, wurde oben bereits erwähnt und in einem Übersichtsartikel erörtert (NEUKAMM & BEYER 2011). Wenn derlei Beispiele das kreative Potenzial von Evolution nicht belegen, was dann?
War Eva ein Klon von Adam?
SCHERER schreibt, JUNKERs wörtliches Verständnis der Schöpfungstexte gehe „sehr weit“, beispielsweise hinsichtlich der Erschaffung Evas aus „der Seite Adams“. In der dazugehörigen Anmerkung 316 fragt er sich dann, ob Eva damit ein Klon Adams gewesen sei, abgesehen davon, dass Gott dafür das Y-Chromosom von Adam entfernt haben müsste (vgl. S. 376)?
Dass JUNKERs Verständnis hier „sehr weit“ gehe, umschreibt mit wohlwollenden Worten, dass diese Interpretation biologisch hanebüchen ist. SCHERER sieht das ganz genau, doch die Konsequenz scheint ihm unangenehm. Somit stellt er lieber eine Frage, statt eine Antwort zu geben.
Eine unverbrämte Antwort hätte wie folgt gelautet: Es ist biologisch ausgeschlossen, dass ein Mann (Geschlechtschromosomensatz = XY) über vegetative Vermehrung (sprich: Klonen) eine Frau (= XX) hervorbringen kann – es sei denn, der „göttliche Designer“ hätte eine Y-zu-X-Chromosomen-Transformation vorgenommen (KUTSCHERA 2014, S. 283 f.). Mal ganz abgesehen davon wäre die Konsequenz gewesen, dass der sowieso schon katastrophal enge Genpool noch weiter eingeengt gewesen wäre, nämlich auf eine einzige Person.
Das Problem mit den Grundtypen
Bei JUNKER lesen wir:
„Erdgeschichtliche Aspekte: Hier geht es um die Frage, ob alle Grundtypen gleichzeitig geschaffen wurden (so verstehe ich das biblische Schöpfungszeugnis) und wie der Fossilbericht verstanden werden kann, der eine teilweise sukzessive Dokumentation von Grundtypen zeigt, die sowohl in Grundtypen- als auch in Evolutionsperspektive als lückenhaft gedeutet wird.“ (S. 354)
Eine Seite später gesteht JUNKER Folgendes ein:
„Die Reihenfolge des fossilen Auftretens von Grundtypen ist dagegen, wie bereits dargelegt, ein ernsthaftes Problem für meine Position.“ (S. 355)
Hintergrund: Kreationisten postulieren evolutionär voneinander getrennte Ursprungseinheiten, sogenannte Grundtypen. Einem Grundtyp sollen sämtliche Organismen angehören, die direkt oder indirekt durch Kreuzungen miteinander verbunden sind. Ein Grundtyp kann taxonomisch eine einzelne Art, eine Gattung oder eine Familie umfassen. So soll es z. B. einen Grundtyp des Menschen (Gattung Homo), einen Grundtyp der Katzen (Familie Felidae) und einen Grundtyp der Finken geben. Innerhalb von Grundtypen werden Artbildungen zugestanden, eine grundtypübergreifende Evolution („Makroevolution“) soll dagegen unmöglich sein. Nach wörtlicher Auslegung der Bibel sollen die einzelnen Grundtypen durch „Schöpfungsakte Gottes“ und „geologisch gesehen ‚gleichzeitig‘ entstanden sein“ (JUNKER & SCHERER 2006, S. 34, 53, 5, 19, 292).
Evolutionsbiologen erwarten vom Fossilbericht im Wesentlichen, dass sich die Fossilien umso mehr von rezenten Formen unterscheiden, je älter sie sind. Sie erwarten beispielsweise Fossilien zu finden, die im Stammbaum „zwischen“ den Sauropsiden und der Kronengruppe der Säugetiere (also auf der Stammlinie der Säugetiere) angesiedelt sind. In der Tat existieren solche Fossilien, etwa in Gestalt basaler Synapsiden, Therapsiden und Cynodontier. Sie dokumentieren die Reihenfolge, in der die Säugermerkmale entstanden sind. Auch der Übergang vom primären zum sekundären Kiefergelenk ist fossil überliefert, der eine bedeutende Transformation auf der Stammlinie der Säugetiere darstellt (vgl. HOSSFELD & JUNKER 2002, S. 176).
Nach dem „Grundtypen“-Konzept dürfte es derartige Fossilien nicht geben. So bemerken JUNKER & SCHERER in ihrem „evolutionskritischen Lehrbuch“:
„Im Rahmen der Schöpfungslehre ergeben sich ebenfalls testbare Schlussfolgerungen: Es wird die Voraussage gemacht, dass unter den Fossilien keine Bindeglieder zwischen Grundtypen gefunden werden." (JUNKER & SCHERER 1998, S. 204)
Zusammenfassung
Liest man die Passagen von JUNKER und SCHERER ohne biologisches und wissenschaftstheoretisches Vorwissen, gewinnt man den Eindruck, die Evolutionsbiologie stecke voller Anomalien und Ungereimtheiten, wogegen es sich bei dem Schluss auf Design um den Schluss auf die beste Erklärung handele. Beides ist nicht der Fall. Die Autoren leiten den Leser in geschickter Weise zum Fehlurteil einer „verminderten Erklärungskraft“ der Evolutionswissenschaften. Vor allem JUNKER gelingt dies dadurch, dass er wichtige Dinge einfach nicht sagt, sich an einem holzschnittartigen Evolutionsbild abarbeitet und Sachverhalte ignoriert, die nicht zu seiner Sicht passen.
Während SCHERER insgesamt vorsichtiger und differenzierter argumentiert, sind JUNKERS Passagen durch Desinformation und karikatureske Zuspitzungen eines Scientific Creationism gekennzeichnet. So gäbe es „systematische (!) Befunde, die auf ein Menschheitsalter von wenigen Tausend Jahren… hinweisen“ (S. 303; Ausrufezeichen im Original). Makroevolution sei aus Sicht heute vorliegender naturwissenschaftlicher Daten „ausgesprochen unglaubwürdig“ (S. 349). Ein „kreatives Potenzial von Evolution“ sei „nicht erkennbar“ (ebd.). Nichtreduzierbar komplexe Strukturen seien „Hürden, die durch Mutation und Selektion nicht genommen werden können“ (S. 141) usw. Derlei Thesen lassen keine kompetente Evolutionskritik erkennen. Dass sein Bild einer kurzen Erdgeschichte mit einem Großteil des naturwissenschaftlichen Hintergrundwissens kollidiert, ist da nur das Tüpfelchen auf dem i.
Unter dem Strich vertritt DROSSEL den wissenschaftlichsten Standpunkt. Sie erkennt die überwältigenden Belege für die evolutive Stammesgeschichte, für eine alte Erde und für eine natürlich ablaufende Evolution. Leider begegnet sie abwegiger Kritik an der Evolutionstheorie oft nur oberflächlich. An einer Stelle verweist sie auf „Diskussionen in den USA“ (S. 342), ohne ein konkretes Argument zu entfalten. So verschenkt sie Argumente, die für die Auseinandersetzung mit dem Kreationismus wertvoll sind.
Nehmen wir die bizarre These zum Beispiel, nichtreduzierbar komplexe Strukturen seien „Hürden, die durch Mutation und Selektion nicht genommen werden können“. DROSSEL hätte ihre Widersacher mit einschlägigen Gegenbeispielen konfrontieren können. Angeboten hätten sich SCHERERs Rückzugsgefecht zur Bakterienflagelle und die Meinungsrevision zur Evolution der eukaryotischen Zelle. Sie hätte die Blutgerinnungskaskade, T-urf13 oder LENSKIs Langzeitexperiment anführen können. Auch der Verweis auf die so wichtige Arbeit von TROTTER et al. (2014) und auf eine Replik zum „Wartezeitproblem“ (vgl. Anmerkung 256 auf S. 305) wären sehr nützlich gewesen. Dass sie diese Möglichkeiten nicht nutzte, ist bedauerlich. Ihre eigene Erwiderung (S. 344 ff.) ist subjektiv und gibt inhaltlich praktisch nichts her.
Spannend wäre ein weiterer Autor aus dem naturalistischen Lager gewesen. Erst dann hätte man von einer ausgewogenen Debatte sprechen können. Das durfte man freilich nicht erwarten, da alle drei Autoren sowie der Verlag, in dem das Buch erschien, ein christlich-apologetisches Interesse verfolgen. Wer glaubt, er könnte mithilfe dieses Buches naturalistischen Positionen adäquat entgegentreten (vgl. S. 347) oder gar vernünftig gegen Evolution argumentieren, der kann nur enttäuscht werden.
Schöpfung und Evolution? Drei Wissenschaftler. Drei Positionen. Eine Debatte
SCM R. Brockhaus, ISBN: 978-3-417-24183-9, Preis: 28,00 €
Fußnote
[1] Bei der „programmierten Variation“ handelt es um die bizarre Vorstellung, dass den ersten Individuen eines Grundtyps (z. B. Adam und Eva) die gesamte jemals mögliche genetische Variabilität und ontogenetische Plastizität einprogrammiert wurde. Damit sollen evolutive Anpassungen ohne weitere schöpferische Eingriffe ermöglicht werden.
Literatur: Siehe Originalartikel
Dieser Artikel wurde zuerst auf der Webseite der AG Evolutionsbiologie veröffentlicht.
Die AG Evolutionsbiologie - Evolution in Wissenschaft, Kultur und Gesellschaft - ist eine gemeinnützige wissenschaftliche Vereinigung. Wir informieren Sie über das interdisziplinäre Konzept der Evolution in Biologie, Kultur und Gesellschaft und geben Einblicke in die moderne Evolutionstheorie und Forschung. Ein weiterer thematischer Schwerpunkt ist die didaktische Aufbereitung und kompetente Zurückweisung der ideologischen Kritik religiös motivierter Evolutionsgegner, um Fehlvorstellungen zur Evolution zu begegnen.
Weitere Informationen auf der Webseite unter der Rubrik »Über uns«.
Kommentare
Neuer Kommentar