Fossilien und Urgeschichte aus biblischer Perspektive von Benjamin Scholl

Bild: Grok
Hinweis: Der Artikel wurde am 19. Oktober 2025 aktualisiert.
Zusammenfassung: Seit jeher faszinieren Dinosaurier vor allem Kinder und Jugendliche. Die Studiengemeinschaft WORT UND WISSEN nutzt dieses Interesse, um gezielt ein junges Publikum anzusprechen. Die farbenfrohe Aufmachung wirkt attraktiv und weckt Neugier. Doch das Buch verfolgt weniger das Ziel, naturwissenschaftlich etabliertes Wissen zu vermitteln. Vielmehr dienen Dinosaurier als Anknüpfungspunkt für eine biblisch-kreationistische Deutung der Urgeschichte. Zwar bezieht sich der Autor auf paläontologische Befunde, doch er verlässt regelmäßig den Bereich der empirischen Nachprüfbarkeit und des wissenschaftlich wohlbegründeten Wissens. Die Befunde werden so gedeutet, dass sie gegen Evolution und die langen geologischen Zeiträume sprechen. So richtet sich das Buch an ein evangelikales Publikum, das etablierte naturwissenschaftliche Erkenntnisse hinterfragen möchte. Eine breitere Wirkung ist angesichts der argumentativen Schwächen jedoch kaum zu erwarten.
Worum geht es in diesem Buch?
Der Autor Benjamin SCHOLL vertritt eine wörtliche Bibelauslegung, was sich in der Auswahl und Darstellung seiner Themen niederschlägt. Im Zentrum stehen unter anderem die Vorstellung einer göttlich intendierten „Programmierung“ der Welt (S. 32), der Tod als Folge des Sündenfalls (S. 52 ff.) sowie die These eines „genialen Designs“ prähistorischer Lebewesen wie der Dinosaurier. Darüber hinaus behandelt er Figuren der biblischen Überlieferung – etwa die Arche Noah (S. 112 ff.) sowie mythische Wesen wie Behemoth und Leviathan (S. 170 ff.).
Einige Aufmerksamkeit widmet er den prähistorischen Ica-Steinen (S. 201), auf denen u. a. Dinosaurier abgebildet sein sollen. Trotz ihrer wissenschaftlich belegten Fälschung rückt er deren Authentizität zumindest in den Bereich des Möglichen und spekuliert, Menschen könnten Dinosaurier durch mündliche Überlieferung gekannt haben.
SCHOLL stützt seine Weltanschauung regelmäßig auf Annahmen, denen es an empirischer und theoretischer Fundierung fehlt. Er verweist z. B. auf Berechnungen zur Schwimmfähigkeit der Arche (S. 116), ohne die Plausibilität einer weltweiten Sintflut zu hinterfragen (S. 118) bzw. in der Sintflut-Erzählung nur einen Mythos zu sehen.
Die Vorstellung, dass sich die enorme Artenvielfalt aus einem extrem reduzierten Genpool innerhalb weniger Jahrtausende entwickelt haben soll, macht die Noah-Geschichte nicht glaubwürdiger. Des Weiteren bemerkt Thomas JUNKER (2009, S. 328):
Da jedes Individuum nur zwei Allele für jedes Gen tragen kann, fragt sich zudem, wo bei Adam und Eva [bzw. den Grundtypen generell; MN] die Vielzahl der historischen, aktuellen und zukünftigen Allele physisch gespeichert war.
Drei von zahlreichen weiteren schwerwiegenden Problemen (vgl. ISAAK 1998) seien hier nur kurz angedeutet:
- Hochseetauglichkeit: Ein Holzschiff von der Größe der Arche (vgl. Abb. 1) ist strukturell instabil, da Holz bei solchen Dimensionen den Kräften von Seegang und Eigengewicht nicht standhält (DUNNING 2011). Nachbauten wie die „Ark Encounter“ mussten mit 95 Tonnen Stahl verstärkt werden und bleiben dennoch an Land. Die Arche des Niederländers Johan HUIBERS wurde mit einer Plattform aus 21 Stahlbargen ausgestattet, um überhaupt schwimmfähig zu sein.
- Umwelt-Schock: Ohne spezialisierte Einrichtungen zur Nachbildung artgerechter Mikroklimata – wie etwa Tropen- und Wüstenhäuser, Polarien, Feuchtbiotope oder Süßwasser-Aquarien – hätten nur wenige Arten auf der Arche überlebt.
- Logistik und Versorgung: Basierend auf der nach WOODMORAPPE (1996) angenommenen Größenverteilung der von ihm geschätzten 16.000 Tiere (bevorzugt Jungtiere) kämen diese nach einem Jahr auf ein Gesamtgewicht von über 400 Tonnen (ISAAK 1998). Diese würden täglich mindestens 30 Tonnen an Nahrung und Frischwasser benötigen und etwa 20 Tonnen Exkremente produzieren.
Selbst wenn die Zahl der Tiere auf 7.700 reduziert würde, wie SCHOLL (S. 119) nach „aktuellem Wissensstand [sic]“ schätzt, von denen ein Teil Winterschlaf hielte (wobei nur wenige Arten physisch dazu in der Lage sind), bliebe der Aufwand exorbitant: Alle Besatzungsmitglieder müssten rund um die Uhr ohne Unterbrechung – bei Regen, Sturm und Dunkelheit – Futter vorbereiten und verteilen, Wasser bereitstellen, die Gehege reinigen, Kot einsammeln und entsorgen, Tiere pflegen und beaufsichtigen und vieles mehr. Ohne technische Hilfsmittel wie Förderbänder, Kräne, Futterspender, Belüftungssysteme etc. wäre das Projekt vollkommen unrealistisch.
Dazu kommt: Viele Arten und ganze Tierfamilien sind extreme Nahrungsspezialisten. Frischer Nektar, lebende Insekten, bestimmte Pflanzen oder Eukalyptusblätter sind oft nur unter definierten Umweltbedingungen und in den natürlichen Habitaten verfügbar. Diese Nahrung ist weder lagerungsfähig noch ersetzbar. Viele Beutegreifer benötigen zwingend Frischfleisch; pflanzliche Ersatzprodukte und Trockenfleisch sind oft unverträglich oder verderblich. Aas wird bereits nach Tagen toxisch. Allein der Heuvorrat für zwei Elefanten wäre gigantisch und würde an der feuchten Luft innerhalb weniger Wochen schimmeln. Ohne moderne Lager-, Kühl-, Konservierungs- und Aufbereitungssysteme ist eine artgerechte Versorgung tausender Tiere über ein Jahr völlig utopisch.

Abb. 1: Die „Wyoming“ war ein hölzerner Sechsmast-Gaffelschoner des 20. Jahrhunderts. Mit einer Rumpflänge von 100 Metern besaß sie den größten hölzernen Segelschiffsrumpf, der je gebaut wurde. Obwohl stahlverstärkt, verzog sich der Rumpf im Seegang. Die Bewegungen bei hohem Wellengang führten zu strukturellen Schäden im Holz, sodass das Schiff permanent leckte. Auf See liefen die Lenzpumpen im Dauerbetrieb, doch weder Stahl noch Pumpen konnten verhindern, dass das Schiff 1924 im Orkan sank. Nun wäre die Arche Noah noch viel länger gewesen – ohne stählerne Stützkonstruktion. Ein seetaugliches Holzschiff dieser Länge, ohne Eisenverstärkungen, wäre technisch extrem unplausibel, wenn nicht unmöglich. Bildquellen: Santa Maria: pxhere, Public Domain. Wyoming: jb.atwood from Huntsville, AL, USA, Schooner Wyoming (26704679510), horizontal gespiegelt, CC BY-SA 2.0. Titanic: HefePine23, Titanic Stardboard Side Diagram, CC BY-SA 2.0. Queen Mary II: Bundesstefan, Queen Mary 2 in a fjord in Norway, CC BY-SA 3.0. Arche Noah: scott1346 from Mechanicsville, MD, USA, Ark encounter (36148737586), CC BY 2.0. Bilder beschnitten und überlagert, Hintergründe entfernt.
.jpg){kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
Wer für das Gelingen eines solchen Unterfangens argumentieren möchte, muss Zusatzannahmen bemühen, die sich unabhängig nicht überprüfen lassen. Er muss auf göttliche Unterstützung (Wunder) setzen, nicht realistische Rahmenbedingungen voraussetzen oder auf ähnlich gewaltiger Holzschiffe aus der nicht belegten Schiffbaugeschichte der Antike verweisen. Mitunter wird etwa die sagenumwobene Tessarakonteres genannt – ein angeblich 130 Meter langes Ruderkriegsschiff aus dem 4. Jahrhundert v. u. Z. (LOVETT 2008). Die umfangreiche Erfahrung der Schiffsbautechnik zeigt jedoch, dass so ein Schiff nie funktioniert hätte (DUNNING 2011).
SCHOLL versucht, auch widerlegende Instanzen in sein biblisches Deutungsraster einzuarbeiten. So wird etwa das Fehlen gemeinsamer Menschen- und Dinosaurierfossilien mithilfe der Mutmaßung wegerklärt,
… dass sich Menschen nach dem Sündenfall von gefährlichen Dinosauriern eher fernhielten – vielleicht hat Gott ihren Lebensraum auch besonders beschützt. (S. 208)
Solche Passagen veranschaulichen, was es bedeutet, „Fossilien und Urgeschichte aus biblischer Perspektive“ zu deuten. Mit Hilfe theologischer Hypothesen über Gottes mögliches Denken und Wirken lassen sich beliebige Befunde mit einem wortwörtlichen Bibelverständnis harmonisieren. So wird der evangelikalen Klientel ein scheinbarer Ausweg aus dem Spannungsverhältnis zwischen moderner Wissenschaft und bibeltreuer Weltdeutung angeboten. Leider geht dies zulasten wissenschaftlicher Kriterien wie empirischer Adäquatheit, Erklärungskraft und externer Konsistenz (Verträglichkeit mit wissenschaftlich gut etabliertem Hintergrundwissen).
Optische Reize und spekulative Darstellungen
Um kreationistische Sichtweisen im Kontext der Darstellung von Dinosauriern spannend und interessant zu gestalten, wird stark auf visuelle Reize gesetzt. Neben knallbunten Textkästchen finden sich leuchtend farbige Abbildungen, die allerdings oft deutlich von paläontologischen Erkenntnissen abweichen (vgl. Abb. 2).

Abb. 2: Links: Darstellung des Styracosaurus bei SCHOLL (S. 84 f.) in leuchtend rot-schwarzer Panzerung. Die gewählte Farbgebung wirkt als Blickfang, ist paläontologisch aber nicht belegt und eine rein künstlerische Interpretation. Plausibler ist eine grün-braune Tarnfärbung (rechts, Public Domain).
{kind=link}
Auch andere Abbildungen zeigen spekulative Merkmale. Auf dem Buchcover sowie auf S. 63 etwa erscheint ein Giganotosaurus mit Rückenstacheln. Ein Tyrannosaurus rex wird auf S. 56/57 in einer phantasievollen blaugrauen Farbgebung mit weißen Flecken dargestellt, im Kampf mit einem Triceratops. Spinosaurus trägt auf S. 187 Stacheln am Kopf, ein weiteres nicht belegtes Merkmal.
Die Bibel als Deutungsrahmen für Anatomie
Neben spekulativen Illustrationen greift SCHOLL zu metaphorischen Deutungen anatomischer Merkmale, die er in den biblischen Kontext überträgt. So wird die Schädeldecke des Pachycephalosaurus (S. 23) als Sinnbild für „Dickköpfigkeit“ gegenüber göttlichen Geboten interpretiert. Der „starre Nacken“ des Triceratops (S. 86 f.) wird wiederum mit Ungehorsam gleichgesetzt, ein in der Bibel oft verwendetes Bild. Solche Analogien sind hermeneutisch fragwürdig und wissenschaftlich nicht belastbar.
Problematisch ist auch SCHOLLs Versuch, Wissenschaften mit historischer Komponente – wie die Paläontologie und Evolutionsbiologie – aus dem Zentralbereich der Naturwissenschaften herauszulösen. Er stellt es so dar, als seien die Ursprünge des Lebens und die Entwicklung der Arten nicht ausschließlich mit naturwissenschaftlichen Methoden zu erforschen (S. 11).
Seine These stützt er auf das „evolutionskritische Lehrbuch“ JUNKER & SCHERER (1998). Darin wird festgestellt, dass historische Begebenheiten weder direkt beobachtbar noch experimentell wiederholbar seien. Dies legt nahe, dass Evolutions- und Schöpfungslehren gleichermaßen auf „weltanschauliche Grenzüberschreitungen“ angewiesen bzw. nicht rein naturwissenschaftlich rekonstruierbar seien. Allerdings ist diese Ansicht wissenschaftstheoretisch nicht haltbar.
Missverständnis historischer Naturwissenschaft
Die Darstellung bei SCHOLL lässt wesentliche Aspekte empirischer Forschung unberücksichtigt. Zunächst: Auch Disziplinen wie Geologie, Evolutionsbiologie, Paläontologie und Kosmologie arbeiten mit theoretischen Modellen, Gesetzesaussagen, Mechanismen, logischen Schlussfolgerungen und (hypothetisch-deduktiver) Bestätigung. Die Modelle werden systematisch überprüft und weiterentwickelt. Wie MAYR (1998) betont, besteht ihre Aufgabe darin, auf Basis aller verfügbaren Daten konsistente und kausal nachvollziehbare Erklärungen zu rekonstruieren.
Vor allen Dingen übersieht SCHOLL, dass nicht die Art der Beobachtung (experimentelle Reproduzierbarkeit, direkte Beobachtbarkeit) eine Naturwissenschaft definiert, sondern der folgerichtige Umgang mit den Daten (MAHNER 1986, S. 41).
So lassen sich zum Beispiel die geologischen Schichten mit archäologischen Siedlungsböden vergleichen: Als Naturprotokoll dokumentieren sie die Entwicklung des Lebens über hunderte Millionen Jahre hinweg. In älteren Schichten finden sich überwiegend einfache, ausgestorbene Organismen; in jüngeren treten stufenweise komplexere, einander abgestuft ähnliche Lebensformen auf, die sich mit abnehmendem Alter zunehmend der heutigen Flora und Fauna annähern.
Da abgestufte Ähnlichkeiten eine logische Folge von gemeinsamer Abstammung, Variation und Selektion sind, liegt der Schluss nahe, dass die Arten eine stammesgeschichtliche Entwicklung durchlaufen haben. Zur Absicherung dieses Schlusses stützt sich die Paläontologie nicht nur auf Fossilien, sondern auch auf Erkenntnisse aus Molekularbiologie, Entwicklungsbiologie, vergleichender Anatomie und Genetik – inklusive unabhängiger Methoden wie molekularer Kalibrierungen.
Ein besonders starker Evolutionsbeleg ergibt sich aus den strukturellen genetischen Markern in unserem Erbgut: Könnten wir in der Zeit zurückreisen und einige Primaten im Miozän mit spezifischen DNA-Markierungen versehen – und würden wir diese Markierungen heute bei Mensch und Schimpanse wiederfinden –, dann wäre ihre gemeinsame Abstammung belegt. Zeitreisen sind unmöglich – doch Retroviren haben genau solche Markierungen vor Millionen Jahren in den Genomen hinterlassen.
Rund 8 % des menschlichen Genoms bestehen aus retroviralen Überbleibseln, sogenannten endogenen Retroviren (ERVs). Ihre virale Herkunft erkennt man an genetischen Strukturelementen, die für Retroviren charakteristisch sind, etwa an den Long Terminal Repeats (LTRs) (vgl. Abb. 3). Die ERVs sehen exakt so aus wie jüngere Einfügungen, die man heute bei Katzen und Schafen kennt. Es gelang sogar, aus menschlichen ERVs eine funktionale Retrovirus-Sequenz zu rekonstruieren, die wieder Viruspartikel bildete (DEWANNIEUX et al. 2006). Ihre virale Herkunft ist damit wissenschaftlich sehr gut abgesichert.
Der springende Punkt: Zahlreiche dieser Markierungen finden sich nicht nur an gleichen Stellen des Erbguts von Mensch, Schimpanse und anderen Primaten, sondern auch in genau der Verteilung, wie sie die Evolutionstheorie erwarten lässt: 384 von 425 vollständigen Retroviren des Schimpansen finden sich beim Menschen an identischen Positionen; zwischen Mensch und Gorilla gibt es weniger Übereinstimmungen, zwischen Mensch und Rhesusaffe noch weniger (POLAVARAPU et al. 2006).
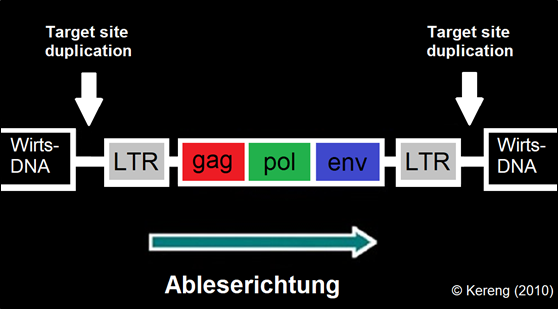
Abb. 3: Schematische Darstellung eines in die Wirts-DNA integrierten ERVs (Provirus). Sie sind Überbleibsel früherer Retroviren-Infektionen, die sich ins Genom der Keimbahn (Ei- oder Samenzellen) integriert haben. Ein Retrovirus besitzt mindestens die drei Gene gag, pol und env: pol kodiert unter anderem die reverse Transkriptase, env kodiert die virale Hülle.
Das Provirus ist an beiden Enden von LTRs (Long Terminal Repeats) begrenzt. Ein typisches Merkmal integrierter Retroviren sind auch die sogenannten Target Site Duplications – kurze, identische DNA-Sequenzen, die bei der Integration des viralen Genoms entstehen und beidseits der Insertionsstelle in der Wirts-DNA erscheinen. Diese Signatur erlaubt es, die Überreste endogener Retroviren im Genom sicher zu identifizieren.
In der Regel werden die Gene epigenetisch stillgelegt oder sind durch Mutationen zerstört – meist sind nur noch die LTRs erhalten. In sehr seltenen Fällen erfüllt ein Gen (niemals aber ein ganzes Provirus) eine nützliche Funktion. Wäre ein ganzes Provirus aktiv, entstünden wieder Viruspartikel – mit potenziell gefährlichen Folgen. Bildquelle: KERENG (2010) / AG Evolutionsbiologie.
Konstruiert man aus der Verteilung der ERVs einen Stammbaum, entspricht er exakt demjenigen, der sich auch aus anderen Merkmalen ableiten lässt. Betrachtet man darüber hinaus jene ERVs, die bei allen Primatenarten an identischer Stelle vorkommen, und analysiert deren Punktmutationen, ergibt sich erneut das schon bekannte stammesgeschichtliche Muster (JOHNSON & COFFIN 1999).
Ob historische Wissenschaft oder nicht – ein derartiger Grad an Bestätigung ist von kaum einer Experimentalwissenschaft zu übertreffen. Dass mehrere voneinander unabhängige Methoden zu übereinstimmenden Ergebnissen gelangen – und zwar genau im Einklang mit den Vorhersagen der Evolutionstheorie – lässt sich kaum vernünftig erklären, wenn diese Theorie falsch wäre.
Kann es eine Schöpfungsforschung geben?
Der Autor verwendet vielfach den Begriff „Schöpfungswissenschaft“ (u. a. auf S. 18 und 69). Dies vermittelt den Eindruck, auf dem Boden biblisch-kreationistischer Vorstellungen und Konzepte könnten wissenschaftliche Disziplinen gedeihen, die valide Erkenntnisse erzeugen. Das Schöpfungsparadigma des Autors lässt sich jedoch nur sehr begrenzt mit jenen Kriterien vereinbaren, denen die empirischen Wissenschaften ihren Erfolg verdanken (vgl. MAHNER 1986, S. 24–38).
Mangels semantischer Klarheit, empirischer Überprüfbarkeit und erklärender Tiefe steht die angenommene Schöpfung erkenntnistheoretisch auf einer Stufe mit teleologischen Konzepten wie der Entelechie. Biblische Vorstellungen im literalen Sinn stehen zudem, wie angedeutet, vor dem grundlegenden Problem, dass sie nicht mit wohletabliertem naturwissenschaftlichem Wissen in Einklang zu bringen sind.
Dem Kreationismus zufolge ist nicht nur die Evolutionstheorie falsch, sondern auch zentrale Erkenntnisse der Anthropologie, Paläontologie, Geowissenschaften, Kosmologie und Planetologie. Selbst elementare physikalische Einsichten wie die Vorgänge in Sternen müssten infrage gestellt werden, da sie die enormen Zeiträume untermauern, die der Kreationismus aus weltanschaulichen Gründen ablehnt. Auch die historisch-kritische Methode der Religionswissenschaft und deren Erkenntnisse werden zurückgewiesen, sofern sie nicht mit der wörtlichen Bibelauslegung in Einklang steht.
Würde man kreationistische Vorstellungen zur Grundlage wissenschaftlicher Erkenntnis machen, bliebe somit vom komplexen, interdisziplinär abgestimmten Theoriengefüge der Naturwissenschaften nur noch ein stark reduzierter Rahmen übrig, dem wesentliche Erklärungszusammenhänge verloren gingen. Vor allem unterminiert die „Schöpfungsforschung“ des Autors die wissenschaftliche Forderung, dass die freie Suche nach Wahrheit nicht durch die Anerkennung von Dogmen oder Autoritäten beeinträchtigt werden darf (MAHNER 2007).
Der Begriff „Schöpfungswissenschaft“ suggeriert also fälschlicherweise Wissenschaftlichkeit, obwohl zentrale notwendige Kriterien nicht erfüllt sind. Bereits in den 1970er-Jahren urteilte ein Gericht in Arkansas, dass die Gleichstellung von „Creation Science“ mit der Evolutionstheorie verfassungswidrig sei, da es sich nicht um empirische Wissenschaft, sondern um religiöse Lehre handle (GEMBALLA & KATTMANN 2024).
Auch die wissenschaftlich gesicherten Details über Dinosaurier stellt SCHOLL häufig nicht korrekt dar. Vieles bewegt sich auf dem Niveau populärer Irrtümer. Dies sei im Folgenden anhand ausgewählter Beispiele erläutert.
Archaeopteryx – nur ein ausgestorbener Vogel?
Kreationisten betrachten Archaeopteryx in der Regel als echten Vogel, da er Federn besaß und zu einfachem Schlagflug fähig war (BOLLEN 2007, S. 222). Evolutionär bedeutsame Übergangsformen existieren für sie nicht, so auch nicht für SCHOLL:
Trotz der Unterschiede zu heute lebenden Vögeln können wir den Archaeopteryx als ausgestorbenen Vogel einordnen, da seine Schlüsselbeine zum sogenannten Gabelbein verschmolzen sind. Zudem hat er eine (seitlich) rückwärts orientierte erste Zehe. (S. 79)
Gabelbeine besaßen aber schon die meisten Nichtvogel-Theropoden, und das bereits seit der Trias (RAUHUT et al. 2020).[1] Eine (seitlich) rückwärts orientierte erste Zehe ist zudem ein Merkmal von Coelurosauriern (OSTROM 1973), das bereits Nichtvogel-Maniraptoren wie Ornitholestes besaßen. Des Weiteren haben neuere Untersuchungen des Archaeopteryx ergeben, dass die Zehenstellung nicht der moderner Vögel, sondern der von Deinonychosauriern entsprach (FOWLER et al. 2011).
Während ein Großteil der Vogelmerkmale auch bei Nicht-Vogel-Dinosauriern auftrat, fehlten dem Archaeopteryx noch viele Merkmale moderner Vögel (KUBI 2025). Umgekehrt besaß der Archaeopteryx noch Dinosaurier-Merkmale, die heutigen Vögeln fehlen. Der mosaikartige Charakter von Archaeopteryx im Übergangsfeld zwischen Nichtvogel-Dinosauriern und Vögeln ist damit klar belegt.
Fehldeutungen: Fasern contra Federn
SCHOLL bezeichnet die von vielen Fachleuten vertretene Einordnung der faserartigen Hautstrukturen bei Flugsauriern als Federn als „Fehlinterpretation“ (S. 45). „Mit Vogelfedern“ habe „das nichts zu tun“. Dabei bleibt unberücksichtigt, dass heutige Vogelfedern spezialisierte Ausprägungen einer Integumentform sind, die sich im Mesozoikum schrittweise zu höherer struktureller Komplexität entwickelte.
Angefangen bei einfachen, haarähnlichen Filamenten über büschelförmige und flaumartige Strukturen bis hin zu voll ausgebildeten Konturfedern sind sämtliche der von Evolutionsbiologen wie PRUM (1999) prognostizierten Übergangsformen der Feder bei nicht-avialen Dinosauriern und frühen Vögeln fossil belegt. Der bei Flugsauriern nachgewiesene Typus repräsentiert die einfachste Form dieser evolutionären Reihe.
Die faserartigen Integumente der Flugsaurier ähneln Vogelfedern in Form, Feinstruktur und chemischer Zusammensetzung, sodass ihre Homologie wahrscheinlich ist (YANG et al. 2018). Ob man sie als „Protofedern“ oder „federhaarartige Integumente“ bezeichnet, ist letztlich eine terminologische Frage.
Falschaussagen über den Fossilbericht
SCHOLL behauptet, „die Vogelmerkmale bei Fossilien in den Gesteinsschichten“ würden „nicht ansatzweise in der Reihenfolge auftreten, wie man sie aus Evolutionsperspektive erwarten würde“ (S. 79). Diese Darstellung ist mit dem aktuellen paläontologischen Erkenntnisstand nicht vereinbar und vermittelt ein Bild, das wesentliche Zusammenhänge der fossilen Befundlage ausklammert.
Zwar gibt es des Öfteren Fossilien mit primitiveren (ursprünglicheren) Merkmalen, die jünger datiert sind als man es auf Basis einer linearen evolutionären Entwicklung erwarten würde. Zum Beispiel zeigt der 130 Mio. Jahre alte Archaeornithura bereits viele Merkmale moderner Vögel, während der 125 Mio. Jahre alte (also jüngere) Confuciusornis noch primitivere Merkmale besaß. Der gleich alte Sinosauropteryx wiederum besaß erst Protofedern.
Zum einen aber zeigt der Fossilienbestand durchaus eine zeitlich gestaffelte Entwicklung von weniger zu stärker vogelähnlichen Merkmalen, was im Einklang mit der Evolutionstheorie steht (vgl. Tabelle 1). Zum anderen ist Evolution kein linearer, sondern ein sich stetig verzweigender Prozess, der viele Seitenlinien hervorbringt, die unterschiedlich lange existierten. Daher sind solche Funde aus Sicht der Evolutionsbiologie erwartbar und spiegeln die Vielfalt und das Nebeneinander von „modernen“ und „ursprünglichen“ Linien wider.
Zusammenfassend lässt sich feststellen, dass sich die Dinosaurierfossilien in eine Reihenfolge bringen lassen, in der sich die Entstehung des Vogelbauplans über eine Serie von Merkmals-Additionen nachvollziehen lässt. Man findet keine Vögel, die sich wesensmäßig von Dinosauriern unterscheiden würden, sondern nur abgestufte Ähnlichkeiten innerhalb der Dinosaurier-Klade.
Dies belegt, dass der Vogelbauplan graduell entstand und dass die Evolution von so charakteristischen Vogelmerkmalen wie Federn und Flügeln nicht von der Selektion auf aerodynamische Eigenschaften hin geprägt war.
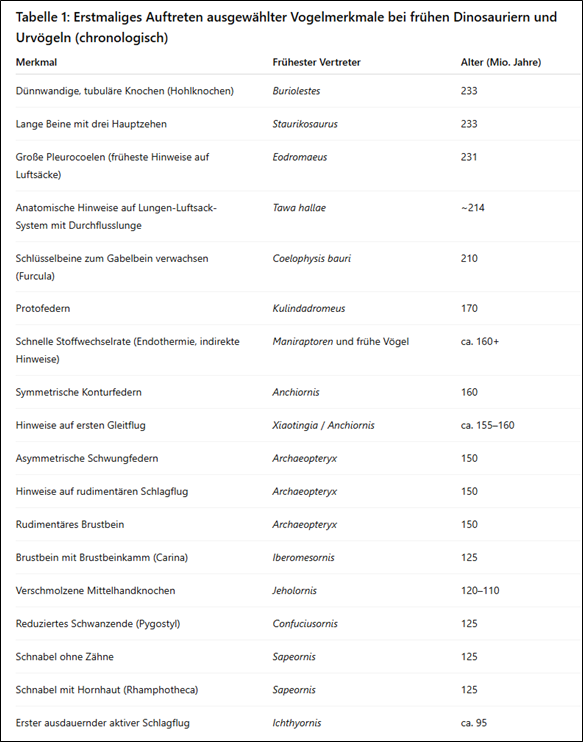
Tabelle 1 ordnet verschiedene vogeltypische Merkmale ihren frühesten bekannten Vertretern und deren Alter in Millionen Jahren zu. Offensichtlich entstanden sie schrittweise und über einen langen Zeitraum hinweg bei frühen Dinosauriern und Urvögeln. Bereits vor über 230 Millionen Jahren traten erste vogelähnliche Merkmale wie hohle Knochen und dreizehige Beine auf, lange bevor es Vögel gab. Komplexere Merkmale wie Federn, Flugfähigkeit oder ein zahnloser Schnabel entwickelten sich deutlich später und in verschiedenen Linien teils unabhängig voneinander.
Gehörknöchelchen der Säugetiere: unwahrscheinliche Konvergenz?
Alle heutigen Säugetiere haben im Innenohr drei Gehörknöchelchen namens Hammer, Amboss und Steigbügel, um den Schall bei Hören zu übertragen und zu verstärken. Die Gehörknöchelchen treten unter den Fossilien so ungleichmäßig verteilt auf, dass Evolutionsbiologen allen Ernstes behaupten, diese seien mehrmals unabhängig voneinander entstanden (= Konvergenzen). Das ist aber sehr unwahrscheinlich! (S. 103)
Diese Aussage ist in zweierlei Hinsicht falsch. Erstens behaupten Evolutionsbiologen keineswegs, dass die Gehörknöchelchen (d. h. die Knochenstrukturen) mehrfach unabhängig entstanden seien. Tatsächlich haben die drei gelenkig miteinander verbundenen Knochen innerhalb der Paukenhöhle (Hammer, Amboss, Steigbügel) in der Abstammungslinie der Mammaliaformes, wie etwa Morganucodon (vgl. Abb. 4), denselben stammesgeschichtlichen Ursprung (siehe z.B. MENG et al. 2011).
Was die Evolutionsbiologen tatsächlich behaupten:
1. Die vollständige Trennung der Gehörknöchelchen vom Kiefer – das heißt ihre Verlagerung und funktionelle Integration ins Mittelohr scheint in der Linie der Monotremen (Kloakentiere) und bei den Theria (Beutel- und Plazentatiere) unabhängig voneinander erfolgt zu sein (RAMÍREZ-CHAVES et al. 2016).
2. Einige Teilstrukturen, wie beispielsweise der Fortsatz des Hammers (das sogenannte Manubrium) scheinen in verschiedenen Linien konvergent entstanden zu sein (MAO et al. 2020). So sehen das Manubrium des Multituberkulaten Sinobaatar pani und das der modernen Säuger ähnlich aus und erfüllen eine ähnliche Funktion. Doch S. pani stammt nicht direkt von denselben Vorfahren wie moderne Säuger ab, bei denen das Manubrium auftritt.
Trotz konvergenter Abkopplung vom Unterkiefer und Verlagerung ins Mittelohr bleiben die Gehörknöchelchen ein gemeinsam abgeleitetes Merkmal der Säugetiere. LUO (2007) spricht daher von der wiederholten Entstehung des „definitiven Mittelohrs“ (DMME), nicht der Gehörknöchelchen selbst.
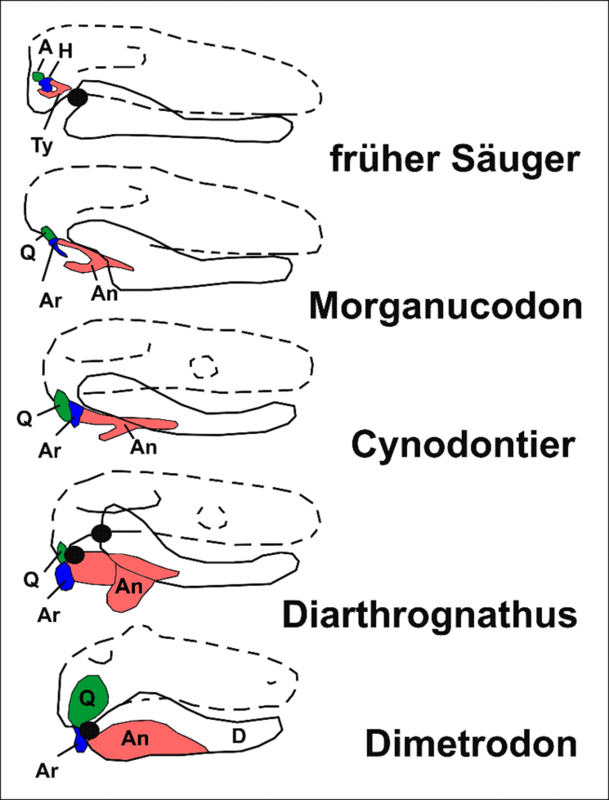
Abb. 4: Die Gehörknöchelchen der Säugetiere stammen von Kieferknochen ursprünglicher Synapsiden wie Dimetrodon ab. Dabei wurde das primäre Kiefergelenk durch ein sekundäres ersetzt – ein Übergang, der ontogenetisch und fossil gut belegt ist (vgl. WANG et al. 2021).
Bei Dimetrodon bildeten Quadratum (Q) und Articulare (Ar) das primäre Kiefergelenk; Angulare (An) und Dentale (D) gehörten zum Unterkiefer. Bei Diarthrognathus sind primäres und sekundäres Kiefergelenk gleichzeitig ausgebildet, das Quadratum bereits stark reduziert. Bei Cynodontiern zeigt das Angulare schon die stimmgabelähnliche Form des späteren Pars tympanica (Ty). Während letzteres bei den Säugetieren Teil des Schläfenbeins ist und mit dem Trommelfell die Paukenhöhle bildet, ist das Mittelohr bei den Cynodontiern noch über den verknöcherten Meckel-Knorpel mit dem Kiefer verbunden.
Kloakentiere, Beuteltiere und Plazentatiere, die sogenannten Kronensäugetiere, haben ein vom Kiefer getrenntes Mittelohr. Bei ihnen ist das Dentale (D) über ein sekundäres Gelenk direkt mit dem Schläfenbein verbunden. Sie besitzen drei Gehörknöchelchen: Hammer (H), Amboss (A) und Steigbügel (nicht im Bild). Hammer und Amboss entstanden aus Articulare bzw. Quadratum, der Steigbügel aus der Columella (hier nicht dargestellt). Entgegen SCHOLLs verkürzender Darstellung vollzog sich nur der letzte Schritt in dieser Transformation, d. h. die Ablösung der Gehörknöchelchen vom Kiefer und ihre Verlagerung ins Mittelohr, mehrfach konvergent.
Der zweite Grund, warum SCHOLL falsch liegt: Konvergente Entwicklungen, also das unabhängige Entstehen ähnlicher Merkmale in unterschiedlichen Linien, ist unter den folgenden Voraussetzungen nicht unwahrscheinlich, sondern hochplausibel:
1. Die wesentlichen Strukturen waren schon vorhanden: Sowohl die „Bauteile“ (Gehörknöchelchen und Muskelansatzstellen) als auch die regulatorischen Mechanismen der Genexpression lagen beim letzten gemeinsamen Vorfahren der Säugetiere als evolutionäres Rohmaterial vor.
2. Ähnliche Selektionsdrücke: Funktionelle Vorteile bestimmter Strukturen – etwa eines Mittelohrs mit abgelösten Gehörknöchelchen zur verbesserten Schallübertragung – begünstigen ähnliche evolutionäre Lösungen.
3. Jeder Teilschritt verbesserte die Hörfunktion: Bereits Articulare und Quadratum leiteten niedrigfrequente Schallwellen über den Steigbügel zur Fenestra ovalis, waren also schon in die Schallübertragung eingebunden (KEMP 2016). Mit der Ausbildung des sekundären Kiefergelenks verkleinerten sich die postdentalen Knochen allmählich, lösten sich vom Kiefer und verlagerten sich schrittweise in Richtung des späteren Mittelohrs (WANG et al. 2021). Dadurch verbesserte sich die Hörfähigkeit, insbesondere durch eine zunehmende Anpassung an das Hören hochfrequenter Töne (LAUTENSCHLAGER et al. 2018).
Fossilien belegen die graduelle Transformation. Der Säuger Liaoconodon hui etwa besaß ein noch wenig spezialisiertes Protomittelohr – eine Übergangsstruktur zwischen frühen Mammaliaformes wie Morganucodon und modernen Säugern: Die inneren Ohrknochen waren teils vom Kiefer getrennt, jedoch noch durch knorpelige und verkalkte Strukturen verbunden, die das Trommelfell stützten.
4. Entwicklungsgene reagieren flexibel: Veränderungen in der Genregulation, etwa im zeitlichen Ablauf (Heterochronie), erklären sowohl die Ablösung der Knochen vom Unterkiefer als auch deren Verlagerung in den Bereich des zukünftigen Mittelohrs.
URBAN et al. (2017) zeigten, dass der TGF-β-Signalweg und eine verstärkte Apoptose maßgeblich am Abbau des Meckel’schen Knorpels beteiligt sind. Modifikationen dieses Signalwegs können diesen Abbau hemmen, was darauf hinweist, dass Veränderungen in der TGF-β-Signalübertragung eine wichtige Rolle bei der evolutionären Umwandlung des Säugermittelohrs gespielt haben.
Konvergenzen, die diese Bedingungen erfüllen, sind also keineswegs ungewöhnlich. Der Dinosaurier, auf den wir im Folgenden zu sprechen kommen, weist eine besonders interessante Kombination konvergenter Merkmale auf.
Chilesaurus: Der Dinosaurier, der nicht ins Schema passt
Dinosaurier werden nach ihrem Becken in zwei große Gruppen eingeteilt: Vogelbecken- und Saurierbecken-Dinosaurier. Allerdings hat der „bizarre“ Chilesaurus aus dem oberen Jura das kräftig durcheinandergewirbelt. Heute streiten Evolutionsbiologen daher heftig, wie man die Dinosaurier überhaupt in einen Stammbaum einordnen soll. (S. 105)
Der Fund des pflanzenfressenden Chilesaurus aus dem Oberjura hat tatsächlich die traditionelle Sichtweise herausgefordert. Chilesaurus besitzt ein ungewöhnliches Mosaik von Merkmalen, die ansonsten ganz unterschiedlichen Dinosauriergruppen zugeordnet werden (vgl. Abb. 5). Deshalb ist seine genaue Position innerhalb des Dinosaurier-Stammbaums schwer bestimmbar.

Abb. 5: Lebendrekonstruktion des Chilesaurus. Da er Merkmale von sowohl Theropoden, Ornithischiern als auch Sauropodomorphen kombiniert, ist seine kladistische Einordnung erschwert. So besitzt Chilesaurus dreizehige Füße wie Theropoden, einen Schnabelansatz wie Ornithischier und einen langgestreckten Hals ähnlich wie Sauropodomorphen. Dies macht ihn zu einer ungewöhnlichen Übergangsform. Welche dieser Merkmale konvergent (unabhängig) entstanden sind, ist bislang nicht abschließend geklärt Bildquelle: Nobu Tamura email: [email protected] http://spinops.blogspot.com/, Chilesaurus NT small, CC BY-SA 4.0.
{kind=link}
Allerdings erwähnt SCHOLL nicht, dass sich auch bei schwer einzuordnenden Arten wie Chilesaurus die meisten ihrer Merkmale eindeutig in die hierarchische Struktur evolutionärer Ähnlichkeiten einordnen. Diese Art von Merkmalshierarchie wird von der Evolutionstheorie vorhergesagt. Angesichts der vorliegenden Befunde ist daher die folgende Schlussfolgerung nicht überzeugend:
Das anhaltende Scheitern, einen stimmigen Stammbaum des Lebens zu erstellen, weist darauf hin, dass er nicht existiert. (S. 105)
Tatsächlich verhält es sich völlig anders: Auch die widersprüchlichen Stammbäume, in denen Chilesaurus unterschiedliche Positionen einnimmt, ähneln sich stark und unterscheiden sich nur in wenigen Details. Merkmalsvergleiche zeigen, dass sich die enorme Zahl theoretisch möglicher Stammbäume reproduzierbar auf eine winzige Teilmenge hochwahrscheinlicher Kandidaten eingrenzen lässt (THEOBALD 2013).
Aus dem Umstand, dass die Rekonstruktion von Verwandtschaftsverhältnissen mitunter schwierig ist, folgt also nicht, dass es keine Stammbäume gibt. Daraus folgt lediglich, dass wir in manchen Fällen noch nicht genau wissen, welcher der relativ wenigen hochwahrscheinlichen Stammbäume der richtige ist.
Die Verwandtschaftsverhältnisse aller wesentlichen Dino-Hauptgruppen sind stabil und unter den allermeisten Experten unstrittig (vgl. RAUHUT & FOTH 2020, S. 37). Trotzdem lassen sich solche Verhältnisse naturgemäß nicht bis in jede Einzelheit rekonstruieren. Der Hauptgrund ist die Unvollständigkeit des Fossilienbefunds: Nur ein äußerst kleiner Teil aller jemals existierenden Arten ist überhaupt fossil überliefert.
Schätzungen zufolge sind weniger als ein bis wenige Prozent aller Arten fossil belegt (RAUP 1994; FOOTE & SEPKOSKI 1999; BENTON et al. 2015; ŽLIOBAITĖ & FORTELIUS 2022). Die Hälfte aller Dinosauriergattungen ist nur durch ein einziges Exemplar belegt; 80 % aller Dinosaurierskelette sind zudem nur fragmentarisch überliefert (DODSON 1990).
Der Leviathan – zwischen Fabel, Fakt und Fantasie
In der biblischen Überlieferung gilt der Leviathan als ein besonders rätselhaftes und faszinierendes „Ungeheuer“. SCHOLL beschreibt ihn als
… das wohl spannendste reale [sic] Tier in der Bibel. Im Buch Hiob heißt es über den Leviathan, dass er Feuer spucken kann. Leider kennen wir bisher kein einziges Wirbeltier, das diese Fähigkeit besitzt. (S. 182)
Im Verlauf seiner Analyse spekuliert SCHOLL über mögliche Kandidaten – etwa den Spinosaurus, verschiedene Krokodilarten oder Plesiosaurier. Sie alle kommen aus verschiedenen Gründen nicht infrage. Da die betreffenden Schilderungen in der Bibel äußerst vage sind, hat die spekulative Zuordnung keine belastbare Grundlage.
In Bezug auf die Fähigkeit des Leviathans, Feuer zu speien, bemüht SCHOLL einen skurrilen Vergleich: Nur der Bombardierkäfer, so meint er, sei zu Ähnlichem fähig. Doch biologisch gesehen ist dieser Vergleich absurd: Bombardierkäfer erzeugen ein chemisches Gemisch, dessen Temperatur etwa 100 °C erreicht. Biologisch sind solche Temperaturen tolerierbar. Die Mundhöhle eines feuerspeienden Wesens hingegen müsste mit hochhitzebeständigen keramischen Materialien ausgekleidet sein, um Temperaturen zwischen 600 °C und 1000 °C zu überstehen. Das ist schlicht Fiktion.
SCHOLL indes verstärkt seine Argumentation dadurch, dass er den einzigartigen Verteidigungsmechanismus des Käfers zum evolutionskritischen Argument erhebt.
Der Bombardierkäfer – evolutionäres Kryptonit aus den ‘70ern
Die 500 bekannten Arten des Bombardierkäfers verfügen über einen hochentwickelten chemischen Abwehrmechanismus (vgl. Abb. 6). Der typische Bombardierkäfer (z. B. Brachinus) erzeugt bei Gefahr in seinem Hinterleib eine reaktive Mischung aus Hydrochinon und Wasserstoffperoxid (H2O2). Im richtigen Moment gibt das Tier die Enzyme Katalase und Peroxidase dazu. Dadurch erhitzt sich die Mischung unter starkem Schäumen bis zum Siedepunkt und tritt explosionsartig aus dem Hinterleib aus. Es entsteht das giftige 1,4-Benzochinon, welches den Fressfeinden unter einem lauten Knall als ätzende, kochend heiße Lösung in Mund und Gesicht spritzt.

Abb. 6: Der Bombardierkäfer schützt sich durch eine chemische Abwehrreaktion, bei der er in seinem Hinterleib zwei Flüssigkeiten (Hydrochinon und Wasserstoffperoxid) mischt. Sobald das Tier bestimmte Enzyme dazu gibt, reagieren sie explosiv miteinander. Dabei entsteht ein heißer, ätzender Spray, den es gezielt auf Fressfeinde schießt, um sie abzuwehren. Illustration: © James Paul BAELLO.
SCHOLL führt an, dass dieser Abwehrmechanismus nichtreduzierbar komplex sei – eine Konstruktion, bei der alle Teile des Abwehrsystems gleichzeitig vorhanden sein müssten, um zu funktionieren. Jede Zwischenform sei funktionslos und daher im Rahmen einer schrittweisen Evolution nicht plausibel erklärbar. SCHOLL wörtlich:
Der Bombardierkäfer ist auch ein explosives Problem für die Evolution: Um eine solche Explosion zu erzeugen, sind nämlich viele verschiedene Komponenten nötig. … Man kann nicht einfach Komponenten wegnehmen, ohne dass die Funktion oder der Schutz in sich zusammenbricht. Das nennt man nichtreduzierbare Komplexität. (S. 183)
Dieses Argument wurde bereits in den 1970er Jahren vorgebracht (z. B. GISH 1977) und ist in der Folge vielfach geprüft und entkräftet worden (etwa WEBER 1981). Inzwischen liegen gut nachvollziehbare Modelle zur Entstehung dieses Mechanismus vor, auf die SCHOLL nicht eingeht.
ISAAK (2007) beispielsweise nennt folgende Evolutionsfolge:
1) Die meisten Insekten produzieren Chinone, etwa zur Sklerotisierung ihrer Kutikula (Außenhülle). Diese stechend riechenden, sehr bitter schmeckenden Substanzen haben den Nebeneffekt, dass sie einige Fressfeinde abschrecken. Verschiedene Käferarten produzieren deshalb verstärkt Chinone und Hydrochinon als Nebenprodukt.
2) Einigen Insektenarten zeigen Spezialisierungen im Körperbau: Vertiefungen oder Drüsen, in denen die Abwehrstoffe gespeichert und ausgeschieden werden können (WEBER 1981). In einem weiteren Schritt wird das Sekret mithilfe von Muskeln abgegeben. Diese Konfiguration existiert bei zahlreichen Käfern, auch bei nahen Verwandten der Bombardierkäfer (FORSYTH 1970).
3) Viele Insekten produzieren reaktive Sauerstoffspezies wie H2O2 – z. B. in der Haut oder im Darm. Die dafür verantwortlichen Enzyme (etwa Glucose-Oxidase) sind uralt. Werden sie in Drüsen exprimiert, führt das zu lokaler H2O2-Produktion. Vermischt sich dieses mit dem bestehenden Wehrsekret, reagiert es mit dem Hydrochinon. Die zunächst noch milde Reaktion beider Komponenten verstärkt die Ungenießbarkeit des Insekts.
4) Werden Peroxidasen, die im Zellstoffwechsel eine wichtige Rolle spielen, verstärkt entlang des Ausführungsgangs gebildet, beschleunigt sich die Reaktion und sorgt für eine erhöhte Konzentration giftiger Chinone im Sekret.
5) Mit zunehmender Enzymaktivität wird die Reaktion intensiver: Sauerstoff wird freigesetzt, und es entsteht ein schaumiges, heißes Gemisch. Dies entspricht einer einfacheren Form des Bombardier-Mechanismus, wie er bei Metrius contractus vorliegt (EISNER et al. 2000).
6) Schlussendlich differenziert sich der Ausführungskanal zu einer verstärkten Reaktionskammer, in der das Sekret vor dem Ausstoß gezielt zur Reaktion gebracht wird. Durch die Optimierung von Enzymproduktion, Kammerstruktur und Ausstoßmechanismus entsteht der effektive „Explosionsapparat“ moderner Bombardierkäfer.
Jeder dieser Schritte ist klein und lässt sich in noch kleinere Teilprozesse aufgliedern. Jede Zwischenstufe ist funktional und teils in verwandten Arten nachgewiesen.
Wo also liegt das „explosive Problem“ tatsächlich? Nicht bei der Evolution, sondern bei dem kreationistischen Mythos! The bombardier beetle myth exploded – so brachte es bereits WEBER (1981) auf den Punkt. Leider scheint kein Argument zu alt und zu abgegriffen, um es nicht gegen die Evolutionstheorie ins Feld zu führen.
Eolithen – menschliche Werkzeuge statt Geröll?
Laut SCHOLL sollen Steinwerkzeuge nicht nur aus den Epochen des Pleistozäns und des oberen Pliozäns stammen, in denen verschiedenste Geräte mit menschlichen Fossilien vergesellschaftet sind. Vielmehr behauptet er, man habe Steinwerkzeuge auch in Schichten entdeckt, in denen es nach vorherrschender Meinung noch keine Menschen gegeben habe:
Die ältesten Steinwerkzeuge wurden sogar in geologischen Schichten des unteren Paläogens (oberes Paläozän) gefunden, das aus evolutionärer Perspektive eine Zeit repräsentiert, zu der es noch nicht einmal Menschenaffen gab. Das ist ein kaum zu überwindendes Problem für die Evolutionstheorie. (S. 210)
Tatsächlich stößt man immer wieder auf Steine, deren Formen und Bruchkanten an (wohlgemerkt: durchweg einfache kantenbestoßene, atypische) steinzeitliche Werkzeuge erinnern. Einige dieser sogenannten Eolithen sind über 50 Millionen Jahre alt.
Allerdings begründete bereits ADRIAN (1948) schlüssig, dass Naturbruchstücke, die wie einfache Schaber, Kratzer, Bohrer usw. aussehen und gedrängt in hochenergetisch verlagerten Sedimenten – etwa in den Grundmoränen eiszeitlicher Gletscher – vorkommen, auf natürliche Weise entstanden sind.
Das Problem: SCHOLL verweist unkritisch auf Michael BRANDTs Buch Vergessene Archäologie. Er unterlässt es jedoch, darauf hinzuweisen, dass diese Steine bereits seit fast 80 Jahren wissenschaftlich als Werkzeuge diskreditiert sind. Eine Auseinandersetzung mit den Argumenten und kritischen Rezensionen zu BRANDTs Werk (etwa NEUKAMM 2017) bleibt aus. Nur auf diese Weise kann SCHOLL eine Aussage vertreten, die aus wissenschaftlicher Sicht seit Jahrzehnten als widerlegt gilt.
Tatsächlich wird BRANDTs Buch von Fachwissenschaftlern nicht als seriöser wissenschaftlicher Beitrag anerkannt, da es an der notwendigen fachlichen Expertise mangelt. Während SCHOLL mit Nachdruck betont, die Formen und Bruchkanten wiesen „viele der ‚Eolithen‘ als echte, menschliche Steinwerkzeuge aus“ (S. 211), weisen Fachleute solche Aussagen aus unbegründet zurück. Ein Beispiel zeigt Abb. 7.
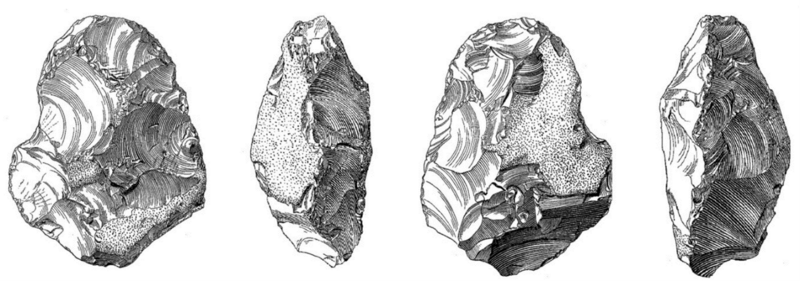
Abb. 7: Eines von vielen Eolithen, die BRANDT als „überzeugende“ menschliche Werkzeuge vorstellt. Handelt es sich hier um einen Millionen Jahre alten Faustkeil? Heutige Experten sind anderer Meinung: Die vermeintliche „Arbeitskante“ zeigt Rinde, die Winkel der Lateralkanten sind stumpf, es fehlt eine Spitze oder umlaufende Schneide, und eine finale Formgebung ist nicht erkennbar. Aus funktionaler Sicht ergibt all dies keinen Sinn. Zudem handelt es sich um ein Stück ohne urgeschichtlichen Bezug. Wer ein solches Stück als artifiziell einstuft, wertet subjektiv. Eine ausführliche Bewertung dieses und anderer Stücke findet sich bei NEUKAMM (2017). Bildquelle: BRANDT (2011, S. 280).
Auch bei Steinen, die BRANDT als besonders aussagekräftig bewirbt, wurde in Gutachten kein artifizieller Charakter festgestellt (vgl. NEUKAMM & HEMMINGER 2020).
Proteine in Dinoknochen als „Zeitanzeiger“
In den letzten Jahrzehnten wurden in Dinosaurierknochen Proteine und Weichteilgewebe entdeckt, was die Diskussion über die mögliche Langzeitkonservierung solcher Materialien neu entfacht hat. SCHOLL nutzt diese Funde, um die langen Erdzeitalter infrage zu stellen und für ein junges Alter der Erde argumentieren. So führt er aus, viele Evolutionsbiologen seien anfangs skeptisch gewesen, da Eiweißstrukturen über Millionen Jahre hinweg nicht erhalten bleiben könnten (S. 214). In dieser vermeintlichen Tatsache sieht er einen Widerspruch zum etablierten Weltalter.
Seine Darlegungen sind aus mehreren Gründen problematisch. Erstens hängt die Zerfallsgeschwindigkeit organischer Materie stark von den Umgebungsbedingungen ab, sodass daraus keine verlässlichen Rückschlüsse auf geologische Zeiträume gezogen werden können. Zweitens liefert die radiometrische Datierung als Goldstandard der Zeitmessung konsistente Ergebnisse, die mit unserem naturwissenschaftlichen Hintergrundwissen übereinstimmen. Und drittens gibt es längst plausible Erklärungsmodelle für die Langzeitkonservierung von Weichgewebe in Fossilien.
Seltsam überzogen wirkt SCHOLLs heroisierende Würdigung der Paläontologin Mary SCHWEITZER, die er als „Wissenschafts-Heldin“ (S. 214) preist – so als habe ihre Arbeit die Grundlagen der geologischen Zeitmessung und Evolutionstheorie erschüttert. Das Gegenteil trifft zu; sie ist eine anerkannte Evolutionsbiologin (und selbst Christin), deren Arbeiten den evolutionären Standpunkt voll untermauern.
SCHWEITZER et al. (2014) zeigen, dass der Abbau des Blutfarbstoffs Hämoglobin Fenton-Reaktionen auslösen kann, welche den langfristigen Erhalt von Gewebe begünstigen. Zudem haben die Kollagen-Reste eines Tyrannosaurus-Exemplars die Bestimmung der Aminosäure-Sequenz und den Abgleich mit der Sequenz bei Vögeln ermöglicht. Dieser Vergleich führt zu genau dem Stammbaum, der zuvor bereits anhand morphologischer Merkmale erstellt wurde (ORGAN et al. 2008).
Im Übrigen würde die Auswertung der Sequenz-Unterschiede über die bekannte Mutationsrate („molekulare Uhr“) Zeiträume liefern, die SCHOLL mit seiner Argumentation gerade in Frage gestellt sehen möchte.
14C-Messungen in Dinosaurierknochen
Nicht nur Weichgewebe im Dinosaurierknochen, auch Radiokohlenstoff (14C oder C14) deutet SCHOLL als Hinweis auf eine vermeintlich junge Erde:
Eine Gruppe von Schöpfungswissenschaftlern veröffentlichte vor einigen Jahren einen interessanten Forschungsbericht: Mit der C14-Methode analysierten sie elf Dinosaurierfossilien, die sie selbst gefunden oder von Museen erhalten hatten… Spätestens nach 50.000 Jahren dürfte kein für uns sicher messbares C14 mehr vorhanden sein. … In den elf untersuchten Dinosaurierfossilien konnten die Wissenschaftler aber C14 finden. …
Achtung – diese Ergebnisse überzeugen nicht alle Schöpfungswissenschaftler! Kritische Stimmen führen an, dass bei anderen, ähnlichen C14-Messungen von Schöpfungswissenschaftlern die Messunsicherheit nicht genügend berücksichtigt wurde. Außerdem könnte es bei den untersuchten Dinoknochen vielleicht Verunreinigungen gegeben haben. …
Wie ich es sehe: Bis das Gegenteil bewiesen wird, sprechen die genannten C14-Messungen in Dinosaurierknochen für mich für eine kurze Erdgeschichte.
Was SCHOLL nicht erwähnt: Fossile Knochen reichern durch verschiedene Prozesse (beispielsweise durch Kontakt mit dem Grundwasser) im Lauf der Jahrtausende sukzessive frisches 14C an (SENTER 2020). Das führt dazu, dass Millionen Jahre alte Fossilien fälschlicherweise als wesentlich jünger erscheinen.
Die Fehlermargen liegt nicht nur im Prozentbereich: Bereits winzigste 14C-Kontaminationen (Ultraspurenbereich) reichen aus, um das Alter eines Millionen Jahre alten Fossils auszuradieren. Wird auch nur 1 ppb (0,0000001%) der ursprünglichen 14C-Menge frisch eingetragen, genügt das, um ein Dinosaurier-Alter von 170.000 Jahren zu erhalten. Radiokohlenstoff-Datierungen von Millionen Jahre alten Proben gestatten daher keine Aussagen über das Alter.
Fazit
Das Buch versucht, die Welt des Mesozoikums in einen bibeltreuen, evangelikalen Deutungsrahmen einzuordnen. Dabei stützt sich der Autor auf Argumente, die in der wissenschaftlichen Fachwelt als problematisch oder widerlegt gelten. Dazu zählen beispielsweise der Hinweis auf angeblich menschliche Werkzeuge aus dem Paläogen (Tertiär) und der Versuch, aus Funden von Weichgewebe in Dinosaurierfossilien Rückschlüsse auf ein junges Erdalter zu ziehen.
Diese Deutungen stehen im Widerspruch zu interdisziplinär bestätigten Erkenntnissen aus Fachbereichen wie Astrophysik, Kosmologie, Geologie, Paläontologie und Evolutionsbiologie. Um seine Sichtweise zu vertreten, muss sie der Autor zurückweisen und Befunde, die sie belegen, entsprechend umdeuten.
Spekulationen über die Separation der Lebensräume von Menschen und Dinosauriern oder über die Konstruktion und Besetzung der Arche verdeutlichen, dass der Autor weniger das Ziel einer strikt wissenschaftlichen Auseinandersetzung verfolgt als die Bestätigung seines bestehenden Weltbildes. Schließlich richtet sich das Buch an ein evangelikal geprägtes, junges Publikum und dessen Eltern. Daher ist zu erwarten, dass die Wirkung des Buchs auf dieses Publikum beschränkt bleibt.
Fußnoten
[1 ] Die Annahme, dass Dinosaurier keine Schlüsselbeine besessen hätten, wurde lange als Argument gegen die Abstammung der Vögel von Dinosauriern angeführt (HEILMANN 1926). Doch bereits 1936 wurden Gabelbeine bei Dinosauriern entdeckt – später auch bei kleinen Theropoden, besonders in der Mongolei (LESSEM 1994, S. 66).
Literatur
Adrian, W. (1948). Die Frage der norddeutschen Eolithen. Schöningh Verlag.
Benton, M. J. (2015). Vertebrate palaeontology (4th ed.). Wiley-Blackwell.
Bollen, L. (2007). Der Flug des Archaeopteryx: Auf der Suche nach dem Ursprung der Vögel. Quelle & Meyer.
Brandt, M. (2011). Vergessene Archäologie. Steinwerkzeuge fast so alt wie Dinosaurier. SCM Hänssler.
Dewannieux, M., Harper, F., Richaud, A., Letzelter, C., Ribet, D., Pierron, G., & Heidmann, T. (2006). Identification of an infectious progenitor for the multiple-copy HERV-K human endogenous retroelements. Genome Research, 16(12), 1548–1556. https://doi.org/10.1101/gr.5565706
Dodson, P. (1990). Counting dinosaurs: how many kinds were there? Proceedings of the National Academy of Sciences, 87, 7608–7612. https://doi.org/10.1073/pnas.87.19.7608
Dunning, B. (2011). Noah's Ark: Sea Trials. Skeptoid Media. https://skeptoid.com/episodes/279
Eisner, T., Aneshansley, D. J., Eisner, M., Attygalle, A. B., Alsop, D. W., & Meinwald, J. (2000). Spray mechanism of the most primitive bombardier beetle (Metrius contractus). Journal of Experimental Biology, 203, 1265–1275.
Foote, M., & Sepkoski, J. J., Jr. (1999). Absolute measures of the completeness of the fossil record. Nature, 398(6726), 415–417. https://doi.org/10.1038/18872
Forsyth, D. J. (1970). The structure of the defence glands of the Cicindelidae, Amphizoidae, and Hygrobiidae (Insecta: Coleoptera). Journal of Zoology, London, 160, 51–69.
Fowler, D. W., Freedman, E. A., Scannella, J. B., & Kambic, R. E. (2011). The predatory ecology of Deinonychus and the origin of flapping in birds. PLoS ONE, 6(12), e28964. https://doi.org/10.1371/journal.pone.0028964
Gemballa, S., & Kattmann, U. (Hrsg.). (2024). Didaktik der Evolutionsbiologie: Zwischen Fachkonzepten und Alltagsvorstellungen vermitteln. Springer Spektrum.
Gish, D. T. (1977). Dinosaurs: Those terrible lizards. Creation-Life Publishers.
Heilmann, G. (1926). The origin of birds. H. F. & G. Witherby.
Isaak, M. (1998). Problems with a global flood (2nd ed.). The Talk Origins Archive. https://www.talkorigins.org/faqs/faq-noahs-ark.html
Isaak, M. (2007). The counter-creationism handbook. University of California Press.
Johnson, W. E., & Coffin, J. M. (1999). Constructing primate phylogenies from ancient retrovirus sequences. Proceedings of the National Academy of Sciences of the United States of America, 96, 10254–10260. https://doi.org/10.1073/pnas.96.18.10254
Junker, R., & Scherer, S. (1998). Evolution: Ein kritisches Lehrbuch (4. Aufl.). Weyel.
Junker, T. (2009). Kreationisten erklären die Evolution: Das „kritische Lehrbuch“ von Reinhard Junker und Siegfried Scherer. In M. Neukamm (Hrsg.), Evolution im Fadenkreuz des Kreationismus. Darwins religiöse Gegner und ihre Argumentation (Religion, Theologie und Naturwissenschaft, Bd. 19, S. 321–338). Vandenhoeck & Ruprecht.
Kemp, T. S. (2016). Non-mammalian synapsids: The beginning of the mammal line. In J. Clack, R. Fay, & A. Popper (Eds.), Evolution of the vertebrate ear (Vol. 59, pp. 107–137). Springer. https://doi.org/10.1007/978-3-319-46661-3_5
Kereng (2010). Evolutionsbeweis durch endogene Retroviren. AG Evolutionsbiologie. https://www.ag-evolutionsbiologie.de/html/2010/evolutionsbeweis-retroviren.html
Kubi, M. (2025). Archaeopteryx: Dinosaurier oder Vogel? https://internet-evoluzzer.de/archaeopteryx-dinosaurier-oder-vogel/
Lautenschlager, S., Gill, P. G., Luo, Z. X., Fagan, M. J., & Rayfield, E. J. (2018). The role of miniaturization in the evolution of the mammalian jaw and middle ear. Nature, 561, 533–537. https://doi.org/10.1038/s41586-018-0521-4
Lessem, D. (1994). Dinosaurierforscher: Die abenteuerliche Suche nach einer untergegangenen Zeit. Birkhäuser.
Lovett, T. (2008). Noah’s Ark. Thinking outside the box. Master Books.
Luo, Z. X. (2007). Transformation and diversification in early mammal evolution. Nature, 450, 1011 1019. https://doi.org/10.1038/nature06277.
Mahner, M. (1986). Kreationismus – Inhalt und Struktur antievolutionistischer Argumentation. Berlin: Pädagogisches Zentrum.
Mahner, M. (2007). Demarcating Science from Non Science. In T. A. F. Kuipers (Hrsg.), Handbook of the Philosophy of Science, Bd. 1: General Philosophy of Science – Focal Issues (S. 515–575). North Holland.
Mao, F., Liu, C., Chase, M. H., Smith, A. K., & Meng, J. (2020). Exploring ancestral phenotypes and evolutionary development of the mammalian middle ear based on Early Cretaceous Jehol mammals. National Science Review, 8, nwaa188. https://doi.org/10.1093/nsr/nwaa188
Mayr, E. (1998). Das ist Biologie: Die Wissenschaft des Lebens. Spektrum Akademischer Verlag.
Meng, J., Wang, Y., & Li, C. (2011). Transitional mammalian middle ear from a new Cretaceous Jehol eutriconodont. Nature, 472, 181–185. https://doi.org/10.1038/nature09921
Neukamm, M. (2017). Vergessene Archäologie: Steinwerkzeuge fast so alt wie Dinos? AG Evolutionsbiologie. https://www.ag-evolutionsbiologie.de/html/2017/vergessene-archaeologie.html
Neukamm, M., & Hemminger, H. (2020). Besprechung: Michael Brandt: Vergessene Archäologie (2. Aufl.). AG Evolutionsbiologie. https://www.ag-evolutionsbiologie.de/pdf/2021/Michael-Brandt-Vergessene-Archaeologie-2020.pdf
Neukamm, M. (2024). Proteine in Dinosaurier-Knochen und der Kreationismus: Warum organische Reste in Fossilien nicht an der geologischen Zeitskala rütteln. AG Evolutionsbiologie. https://www.ag evolutionsbiologie.de/html/2024/proteine fossilien knochen kreationismus.html
Organ, C. L., Schweitzer, M. H., Zheng, W., Freimark, L. M., Cantley, L. C., & Asara, J. M. (2008). Molecular phylogenetics of Mastodon and Tyrannosaurus rex. Science, 320, 499. https://doi.org/10.1126/science.1154284
Ostrom, J. H. (1973). The ancestry of birds. Nature, 242, 136. https://doi.org/10.1038/242136a0
Prum, R. O. (1999). Development and evolutionary origin of feathers. Journal of Experimental Zoology, 285, 291–306. https://prumlab.yale.edu/sites/default/files/prum_1999_mde_development.pdf
Ramírez Chaves, H. E., Weisbecker, V., Wroe, S., & Phillips, M. J. (2016). Resolving the evolution of the mammalian middle ear using Bayesian inference. Frontiers in Zoology, 13, Article 39. https://doi.org/10.1186/s12983 016 0171 z
Rauhut, O. W. M., & Foth, C. (2020). The origin of birds: current consensus, controversy, and the occurrence of feathers. In Foth, C., & Rauhut, O. (Eds.) The evolution of feathers. From their origin to the present (S. 27–45). Springer Nature.
Rauhut, O. W. M., Foth, C., & Tischlinger, H. (2020). Archaeopteryx und andere Urvögel aus dem Solnhofener Archipel. Archaeopteryx, 36, 4–15.
https://www.researchgate.net/publication/342746261_Archaeopteryx_und_andere_Urvogel_aus_dem_Solnhofener_Archipel
Raup, D. M. (1994). The role of extinction in evolution. Proceedings of the National Academy of Science, 91, 6758–6763. https://doi.org/10.1073/pnas.91.15.6758
Polavarapu, N., Bowen, N. J., & McDonald, J. F. (2006). Identification, characterization and comparative genomics of chimpanzee endogenous retroviruses. Genome Biology, 7(6), R51. https://doi.org/10.1186/gb-2006-7-6-r51
Schweitzer, M. H., Zheng, W., Cleland, T. P., Goodwin, M. B., Boatman, E., Theil, E., …, & Fakra, S. C. (2014). A role for iron and oxygen chemistry in preserving soft tissues, cells, and molecules from deep time. Proceedings of the Royal Society B: Biological Sciences, 281(1775), 20132741. https://doi.org/10.1098/rspb.2013.2741
Senter, P. J. (2020). Radiocarbon in dinosaur fossils: Compatibility with an age of millions of years. The American Biology Teacher, 82(2), 72–79. https://doi.org/10.1525/abt.2020.82.2.72
Theobald, D. (2013). 29+ Evidences for macroevolution. Part 1: The unique universal phylogenetic tree. https://www.tinyurl.com/4kzfp5jt
Urban, D. J., Anthwal, N., Luo, Z. X., Maier, J. A., Sadier, A., Tucker, A. S., & Sears, K. E. (2017). A new developmental mechanism for the separation of the mammalian middle ear ossicles from the jaw. Proceedings of the Royal Society B: Biological Sciences, 284, Article 20162416. https://doi.org/10.1098/rspb.2016.2416
Wang, J., Wible, J. R., Guo, B., Shelley, S. L., Hu, H., & Bi, S. (2021). A monotreme like auditory apparatus in a Middle Jurassic haramiyidan. Nature, 590, 279–283. https://doi.org/10.1038/s41586-020-03137-z
Weber, C. G. (1981). The bombardier beetle myth exploded. Creation/Evolution, 2(1), 1–5. https://ncse.ngo/bombardier-beetle-myth-exploded
Woodmorappe, J. (1996). Noah's ark: A feasibility study. Institute for Creation Research.
Yang, Z., Jiang, B., McNamara, M. E., Kearns, S. L., Pittman, M., Kaye, T. G., …, & Benton, M. J. (2018). Pterosaur integumentary structures with complex feather-like branching. Nature Ecology & Evolution, 3, 24–30. https://doi.org/10.1038/s41559-018-0728-7
Žliobaitė, I., & Fortelius, M. (2022). On calibrating the completometer for the mammalian fossil record. Paleobiology, 48(1), 1–11. https://doi.org/10.1017/pab.2021.22
Dinosaurier und die Bibel: Fossilien und Urgeschichte aus biblischer Perspektive
Christl. Verlagsgesellschaft Dillenburg, 2025

Kommentare
In meiner Rezension der ersten Auflage (Kurzfassung Waschke, T. (2025) 'Benjamin Scholl: Dinosaurier und die Bibel' Skeptiker 2025 (1):44, Langfassung als Rezension bei amazon) habe ich weniger Wert auf die fachwissenschaftlichen Fehldarstellungen des Autors gelegt, sondern versucht, zu zeigen, dass dieses Buch ein Kompendium dessen darstellt, was als Kurzzeit-Kreationismus vor allem den USA seit den 1960er Jahren eine Art Standard der Evolutionsgegner geworden ist. Je nach Umfrage bekennen sich fast 50 Prozent der Amerikaner zu derartigen Auffassungen (die Erde ist jung, es gab eine weltweite Sintflut und eine Schöpfungswoche). Die Rezension von Klaus Steiner und meine ergänzen sich daher.
Scholl hat mit diesem Buch die meines Wissens aktuellste und umfassendste Darstellung dieses Weltbilds aus deutscher Sicht (im Gegensatz zu dem US-Amerikaner vertritt die Studengemeinschaft Wort und Wissen den Kurzzeit-Kreationismus nicht als bessere Wissenschaft, sondern aus Glaubensgründen entgegen vielen aktuellen Erkenntnissen, aus Sicht der US-Amerikaner sind sie daher 'Weicheier', aus diesem Grund sind so gut wie keine Arbeiten der deutschen Kreationisten ins Englische übersetzt worden, mit Ausnahme einiger Arbeiten von Werner Gitt) vorgelegt.
Das Buch sollte jeder gelesen haben, der sich für Kurzzeit-Kreationismus interessiert oder aus sich aus irgendwelchen Gründen mit dieser Pseudowissenschaft befassen muss. Man muss sich dazu das Buch nicht kaufen, das erste Kapitel kann man kostenlos unter
https://www.wort-und-wissen.org/produkt/dinosaurier-bibel
(Link ganz unten, nach einer allgemeinen Beschreibung des Buchs).
Antworten
Hallo Herr Waschke,
dass Sie in Ihrer Rezension "weniger Wert auf fachwissenschaftliche Fehldarstellungen des Autors gelegt haben", lässt mich etwas verwundert zurück. Es klingt ein wenig wie ein Eingeständnis, dass Ihnen gewisse Fehler nicht aufgefallen sind...?
Meiner Meinung muss man das Buch überhaupt nicht gelesen haben; es ist entbehrlich, sich mit Pseudowissenschaft auseinanderzusetzen. Da macht es doch mehr Sinn, wenn man sich ein Fachbuch zulegt - und genau das versuche ich in meiner Rezension aufzuzeigen!
Antworten
Neuer Kommentar